|
<NOTE>
Published online (June 16, 2020)
Nkala Forest: Introduction of a forest-savanna mosaic field site of wild bonobos and its future prospects
Ena Onishi1
1 Wildlife Research Center, Kyoto University, Japan 2 Mbou-Mon-Tour, Democratic Republic of the Congo 3 Japan Monkey Centre, Japan 4 Primate Research Institute, Kyoto University, Japan 5 Institute for Advanced Study, Kyoto University, Japan INTRODUCTION Bonobos are one of humans two closest living relatives and display several remarkable similarities to us (Hare & Yamamoto 2015; 2017). To understand human evolution, it is essential to understand bonobos. However, compared to chimpanzees, research on bonobos is lacking. There are far fewer bonobo research sites, with far less environmental diversity. In this article, we introduce our wild bonobo research site located at the southwest end of bonobos range in the Forest Concession of Local Communities of the Mbali River (hereafter, Mbali area, previously called Malebo), in Lac Tumba landscape, Mai-Ndombe province, Democratic Republic of the Congo (Figure 1). The site is unique for its forest-savanna mosaic environment, as previous studies on wild bonobos have mainly been conducted in rich tropical rainforests (Furuichi & Thompson 2010) such as Wamba (Kano 1992), Lomako (Badrian & Malenky 1984) and LuiKotale (Hohmann & Fruth 2003). Inogwabini et al. (2008) recently confirmed that wild bonobos inhabit a wider range of environments than previously thought, and since then much attention has been paid to wild bonobos in forest-savanna mosaic environments (Inogwabini et al. 2013; Narat et al. 2015a, b, c; Pennec et al. 2016; Serckx et al. 2014, 2015, 2016). Wild bonobos in Mbali thus present a promising and novel research opportunity for understanding bonobo behavior from a broader species level. Studies in forest-savanna mosaic environment are also important for theories of human evolution. Many researchers suggest that much of human evolution occurred in forest-savanna mosaic environments after divergence from a forest-dwelling ancestor, and it is therefore essential to study how savanna environments influence great apes behavior and society. Such an approach has been taken to some extent with chimpanzees (e.g., Pruetz & Bertolani 2009), however there are few studies with wild bonobos. Some research in forest-savanna mosaic environments has been conducted on local ecology (e.g., Serckx et al. 2015; Trolliet et al. 2016; Pennec et al. 2016) and human-bonobo relationships (Inogwabini et al. 2013; Narat et al. 2015a) in the Mbali area, and also on conservation (Hickey et al. 2013) and genetics (Kawamoto et al. 2013; Takemoto et al. 2017) in Tshuapa-Lomami-Lualaba (TL2) in the eastern end of bonobos range (Figure 1). However, studies on bonobos behavior and society in forest-savanna mosaic environments are extremely limited (but see Druelle et al. 2020; Narat et al. 2015b). 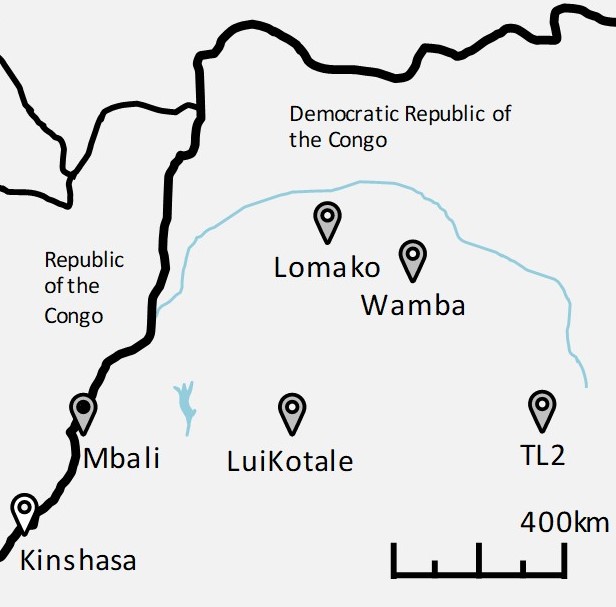
Figure 1. Map of bonobo research sites listed in this paper. The Nkala is one of the field sites in Mbali which is located in the Lac Tumba landscape. GENERAL INFORMATION Mbou-Mon-Tour (MMT), a local NGO for conservation, eco-tourism, and community development, started habituating wild bonobos in the Nkala, Mpelu and Manzano sites in 2001. The first two sites have been technically and financially supported by WWF-DRC since 2006, and the third site, Manzano, has been supported by Bonobo-ECO since 2010. MMT has been operating daily monitoring of bonobos from their morning to night nests whenever possible. Since 2014, in collaboration with MMT and WWF-DRC, Kyoto University researchers have studied the Nkala group (named after the nearest village, 2°36'10"S, 16°27'17"E). Thanks to collaborative efforts of extensive habituation, we have been able to identify all bonobos in the Nkala group since 2016 and can observe them from a distance of approximately 10 to 20 m. This area includes old and young secondary forests, savanna, crop fields and villages (Figure 2). In the Nkala forest, we found direct or indirect evidence for 44 species of mammal including pangolins, civets, duikers, bonobos, and other primates such as Cercopithecus ascanius and Cercopithecus neglectus (Shintaku et al. unpublished data). Direct evidence included live observation, hearing vocalizations and camera trap footage, while indirect evidence included hair, feces, footprints, and reports from the local people. 
Figure 2. Aerial photo of Nkala forest taken by a drone. (Photo taken by Ena Onishi) COMPOSITION OF THE BONOBO GROUP The Nkala group consisted of around 15 individuals until a drastic population decline in 2018. Table 1 shows the number of confirmed individuals at the end of each year from 2016 to 2019. In 2017, two females disappeared. An adult female was suspected to have died from natural causes and a juvenile female was suspected to have transferred to another group. In 2018, four adult and subadult males disappeared; one adult male disappeared in late March, two adult males disappeared simultaneously in early June, and another subadult male disappeared in late June. MMT research assistants did not detect any signs of disease or injury in their daily follows, and the infants survived, suggesting a disease epidemic is unlikely. We did not find dead bodies of bonobos during the study period. At the end of 2019, the population had only 10 individuals, including 4 juveniles and infants. 
HABITAT USE Since 2016, in collaboration with MMT local assistants, we have recorded bonobos activity and habitat use every 30 minutes on full day follows whenever possible. We analyzed their habitat use throughout 2018; 280 days of data were collected (23.3±5.0 days in a month on average) including 6997 data points. During observation, research assistants recorded bonobos location with four categories: closed canopy forest, Marantaceae forest, inundated forest, and savanna (Tutin et al. 1994). Marantaceae forest refers to a forest type where Marantaceae plants predominate the landscape as shown in Figure 3. 
Figure 3. Marantaceae forest. (Photo taken by Ena Onishi) Figure 4 shows monthly habitat use by the Nkala group. They spent 47.9% of the total time in closed canopy forest, 50.2% in Marantaceae forest, 1.4% in inundated forest, and 0.5% in savanna environments. There was a significant difference in habitat use between the dry and wet season (dry season: February to March and June to August; wet season: April to May and September to January (Inogwabini et al. 2008); Fishers exact test: p-value < 0.01). Multiple comparison with Holm correction revealed that bonobos significantly increased the ratio of savanna use in wet season (0.18% in dry season; 0.85% in wet season). 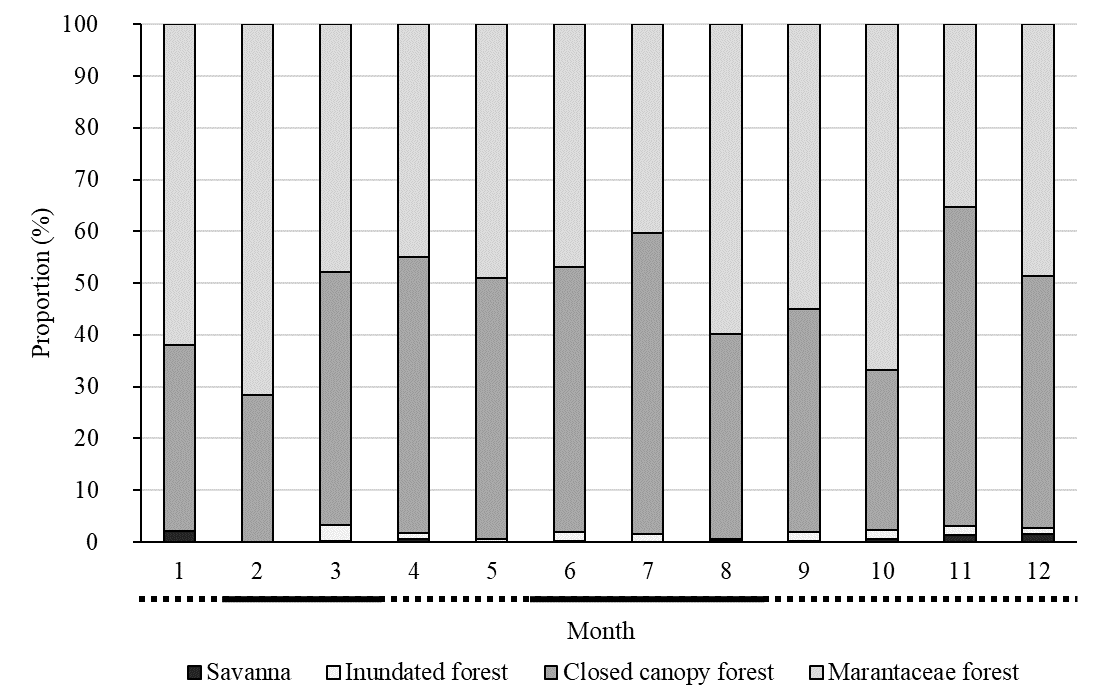
Figure 4. Habitat use by the Nkala bonobos in 2018. The black line along the x-axis represents dry season and the dotted line represents wet season. Crossing to a distant forest was the most common use of savanna in our observations (see video 1). We observed four complete savanna crossing events, each spanned less than 10 minutes. We have also observed longer stays (more than 2 hours) and foraging behavior in the savanna (supplementary video 2; we could not identify the food item in the video). CONSERVATION Nkala villagers (Bateke people) have a strong traditional taboo against eating bonobos (Inogwabini et al. 2013). Due to the MMTs long-term efforts in the Mbali area (e.g. Narat et al. 2015b), the Nkala forest is protected by the community. Hunting of bonobos is strictly prohibited, though hunting of small animals with traditional methods is permitted. Unfortunately, during their regular patrolling of the forest twice a month, MMT forest rangers have found numerous banned metal-wire traps and cartridges. They found 38 traps and 57 cartridges between July 2018 and May 2019 (Table 2). Suspected poachers with guns were also recorded by our camera traps. 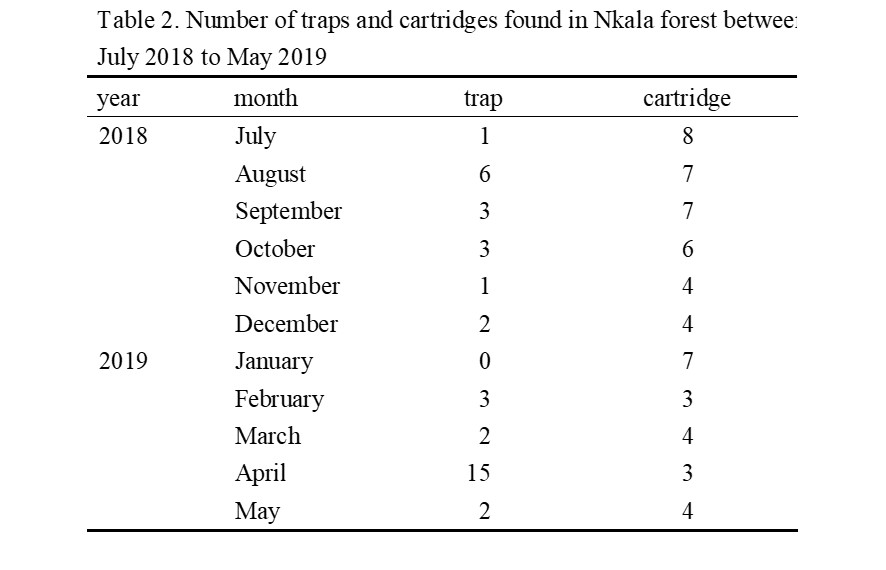
DISCUSSION Our preliminary survey and analyses on the Nkala bonobos habitat use shows constant and strong preference for forest and Marantaceae forest over inundated forest or savanna. Bonobos spent just 02.1% of time in savanna, despite the fact that 40% of their habitat by area is savanna (Inogwabini et al. 2008), suggesting that bonobos constantly and strongly avoid use of savanna. This is unsurprising considering the savanna has greater visibility, lower food availability and increased human activity. However, based on our observations, we can also predict that bonobos forage for plants or insects in the savanna which are not available or rare in the other habitat categories. This perspective is supported by the differential time spent in the savanna between dry and wet seasons, though this should be investigated further. Future research, especially cross-site comparisons, can help reveal their adaptation to forest-savanna mosaic environments, which may prove insightful for understanding human evolution in similar environments. Despite the evident scientific value of this field site, the Nkala group is at a critical risk of extinction. The traps and cartridges found in the forest indicate it is possible that the decline in group size was due to illegal poaching, though there is no direct evidence of bonobo hunting. Poaching for the commercial bushmeat trade is one of bonobos main threats (IUCN & ICCN 2012), and the situation in Nkala may be especially critical because the forest is directly beside a village. Thus, community development is key for conservation, and establishment of a sustainable economic alternatives to hunting activities is essential. Eco-tourism may be one of the most effective and sustainable strategies, thanks in part to the sites close proximity to the countrys capital, Kinshasa (1-hour flight or 1-day boat trip). For this purpose, an international presence at this site must be established to attract tourists. MMT has detected several other bonobo groups around the Nkala forest besides the Manzano and Mpelu groups, but intergroup relationships and migration patterns are not yet known. Green-corridor planting projects like those at Bossou-Nimba, Guinea (Matsuzawa et al. 2011) might help ensure connectivity and migration between forest fragments despite savanna, crop and grazing fields, and roads separating them. As scientific researchers, we would like to continue our efforts to study the wild bonobos in this area, promote their scientific importance, and acknowledge and support the local community that has made tremendous efforts to conserve the forest and the bonobos. We hope this report can help raise awareness of the conservation issues and the research possibilities in Nkala. ACKNOWLEDGMENTS We thank Mbou-Mon-Tour for their support and permission to conduct our research at the field site. We also thank Prof. Raymond Lumbuenamo of World Bank, ERAIFT, and WWF-DRC for their continuous collaboration in this project. Support from the local community at Nkala is also greatly appreciated. We are also grateful to Dr. Victor Narat for his valuable comments on our earlier version of this manuscript. This research was financially supported by the Leading Graduate Program in Primatology and Wildlife Science of Kyoto University, and JSPS grant-in-aid for scientific research (KAKENHI 15H05309 and 19H00629 to Shinya Yamamoto). REFERENCES Badrian N, Malenky RK 1984. Feeding ecology of Pan paniscus in the Lomako forest, Zaire. In: The Pygmy Chimpanzee. Susman RL (eds), Springer, Boston, pp. 275299. https://doi.org/10.1007/978-1-4757-0082-4_11 Druelle F, Aerts P, Bokika JC et al. 2020. Impressive arboreal gap-crossing behaviors in wild bonobos, Pan paniscus. Int J Primatol 41: 129140. https://doi.org/10.1007/s10764-020-00140-z Furuichi T, Thompson J 2010. The Bonobos: Behavior, Ecology, and Conservation. Springer, New York. https://doi.org/10.1007/978-0-387-74787-3 Hare B, Yamamoto S 2015. Moving bonobos off the scientifically endangered list. Behaviour 152: 247258. https://doi.org/10.1163/1568539X-00003263 Hare B, Yamamoto S 2017. Bonobos: Unique in Mind, Brain, and Behavior. Oxford University Press, Oxford. https://doi.org/10.1093/oso/9780198728511.001.0001 Hickey JR, Nackoney J, Nibbelink NP et al. 2013. Human proximity and habitat fragmentation are key drivers of the rangewide bonobo distribution. Biodivers Conserv 22: 30853104. https://doi.org/10.1007/s10531-013-0572-7 Hohmann G, Fruth B. 2003. Lui Kotal: A new site for field research on bonobos in the Salonga National Park. Pan Afr News 10: 2527. https://doi.org/10.5134/143430 IUCN & ICCN 2012. Bonobo (Pan paniscus): Conservation Strategy 20122022. IUCN/SSC Primate Specialist Group & Institut Congolais pour la Conservation de la Nature, Gland. Inogwabini BI, Bewa M, Longwango M et al. 2008. The bonobos of the Lake Tumba - Lake Maindombe hinterland: Threats and opportunities for population conservation. In: The Bonobos: Behavior, Ecology, and Conservation. Furuichi T, Thompson J (eds), Springer, New York, pp. 273290. https://doi.org/10.1007/978-0-387-74787-3_14 Inogwabini BI, Nzala AB, Bokika JC 2013. People and bonobos in the southern Lake Tumba landscape, Democratic Republic of Congo. Am J Hum Ecol 2: 4453. https://doi.org/10.11634/216796221302309 Kano T 1992. The Last Ape: Pygmy Chimpanzee Behavior and Ecology. Stanford University Press, Stanford. Kawamoto Y, Takemoto H, Higuchi S et al. 2013. Genetic structure of wild bonobo populations: diversity of mitochondrial DNA and geographical distribution. PLoS One 8: 19. https://doi.org/10.1371%2Fjournal.pone.0059660 Matsuzawa T, Ohashi G, Humle T et al. 2011. Green corridor project: Planting trees in the savanna between Bossou and Nimba. In: The Chimpanzees of Bossou and Nimba. Matsuzawa T, Humle T, Sugiyama Y (eds), Primatology Monographs. Springer, Tokyo, pp.361370. https://doi.org/10.1007/978-4-431-53921-6_38 Narat V, Guillot J, Pennec F et al. 2015a. Intestinal helminths of wild bonobos in forest-savanna mosaic: Risk assessment of cross-species transmission with local people in the Democratic Republic of the Congo. Ecohealth 12: 621633. https://doi.org/10.1007/s10393-015-1058-8 Narat V, Pennec F, Bisson, LL et al. 2015b. Bonobo conservation as a means for local development: An innovative local initiative of community-based conservation in Democratic Republic of Congo. J Primatol 4: 100127. Narat V, Pennec F, Simmen B et al. 2015c. Bonobo habituation in a forestsavanna mosaic habitat: Influence of ape species, habitat type, and sociocultural context. Primates 56: 339349. https://doi.org/10.1007/s10329-015-0476-0 Pennec F, Krief S, Hladik A et al. 2016. Floristic and structural vegetation typology of bonobo habitats in a forest-savanna mosaic (Bolobo territory, D.R.Congo). Plant Ecol Evol 149: 199215. https://doi.org/10.5091/plecevo.2016.1157 Pruetz JD, Bertolani P 2009. Chimpanzee (Pan troglodytes verus) behavioral responses to stresses associated with living in a savanna-mosaic environment: Implications for hominin adaptations to open habitats. PaleoAnthropology 2009: 252262. https://doi.org/10.4207/PA.2009.ART33 Serckx A, Huynen MC, Bastin JF et al. 2014. Nest grouping patterns of bonobos (Pan paniscus) in relation to fruit availability in a forest-savannah mosaic. PLoS ONE 9: e93742. https://doi.org/10.1371/journal.pone.0093742 Serckx A, Huynen MC, Beudels-Jamar RC et al. 2016. Bonobo nest site selection and the importance of predictor scales in primate ecology. Am J Primatol 78: 13261343. https://doi.org/10.1002/ajp.22585 Serckx A, Kühl HS, Beudels-Jamar RC et al. 2015. Feeding ecology of bonobos living in forest-savannah mosaics: Diet seasonal variation and importance of fallback foods. Am J Primatol 77: 948962. https://doi.org/10.1002/ajp.22425 Takemoto H, Kawamoto Y, Higuchi S et al. 2017. The mitochondrial ancestor of bonobos and the origin of their major haplogroups. PLoS One 12: 114. https://doi.org/10.1371/journal.pone.0174851 Trolliet F, Serckx A, Forget PM et al. 2016. Ecosystem services provided by a large endangered primate in a forest-savanna mosaic landscape. Biol Conserv 203: 5566. https://doi.org/10.1016/j.biocon.2016.08.025 Tutin CEG, White LJT, Williamson EA et al. 1994. List of plant species identified in the Northern Part of the Lopé Reserve, Gabon. Tropics 3: 249276. https://doi.org/10.3759/tropics.3.249 Received: 28 April 2020 Accepted: 1 June 2020 Back to Contents |