|
<NOTE>
Termite Fishing by Mahale Chimpanzees: Revisited, Decades Later
Alejandra Pascual-Garrido
School of Archaeology, University of Oxford, UK INTRODUCTION Jane Goodalls discovery in 1960 that wild chimpanzees at Gombe Stream National Park in Tanzania regularly make and use tools to fish for termites was of great importance, as scientists thought at the time that humans were the only species to make and use tools (Goodall 1964). On hearing of Goodalls observation, her mentor, the paleoanthropologist Louis Leakey, famously remarked: “Now we must redefine ‘tool’, redefine ‘man’, or accept chimpanzees as ‘humans’”. But did only Gombe chimpanzees do this? Since then, evidence of termite fishing has been described in many wild populations of chimpanzees, and is now recognized as one of the most widespread forms of chimpanzee technology (McGrew 1992). The first evidence of termite fishing by the chimpanzees of Bilenge (B group) that inhabit Mahale Mountains National Park in Tanzania dates to 1975. At that time, a local field assistant to the Japanese team of primate researchers by chance discovered recently abandoned plant tools close to a termite mound (Nishida & Uehara 1980). In the following years, more ad libitum records followed. These findings suggested that chimpanzees of B group habitually fish for termites, using 16 plant species to manufacture implements made (mostly) from bark, but also from sedges, vines, twigs and leaves (Nishida & Uehara 1980; McGrew & Collins 1985). Over the last three decades, termite fishing of B group has remained unstudied, as research focus shifted to the habituated neighboring Mimikire (M) group (Nakamura et al. 2013). However, my recent research provides evidence that the Bilenge chimpanzees have continued with their termite fishing tradition, providing yet another example of the long-term maintenance of material culture traditions in our closest living relatives. METHODS The unhabituated chimpanzees (Pan troglodytes schweinfurthii) of B group live around Bilenge (S 6°2′, E 29°44′; 772–1550 m altitude)in the Mahale Mountains National Park, western Tanzania (Nishida 1968; McGrew & Collins 1985). Bilenge is open grassy woodland, mostly miombo, with narrow strips of vine tangle and valley forests with broad hilltops rising from the coastal plain (Collins & McGrew 1988). Earlier studies indicated that termite fishing at Bilenge is seasonal, with the most productive months in November and December coinciding with the annual reproductive and dispersal cycles of the termites (McGrew & Collins 1985). My fieldwork, aided by Tanzanian field assistants, thus encompassed three such periods (25 Nov–14 Dec 2014; 20 Oct–8 Nov 2015; 21 Nov–8 Dec 2016). During reconnaissance surveys, we first searched for mounds that had been targeted previously by chimpanzees, defined by one or more of the following criteria (cf., Pascual-Garrido et al. 2012; Almeida-Warren et al. 2017): (a) tools, fragments or modified raw materials were present; (b) nearby plants had been sourced for raw material, indicated by, e.g., broken stems or branches or parts removed; (c) abandoned tools could be retrospectively fitted to their sources. Measurements included tool characteristics such as length, diameter, type of raw material, approximate age (estimated number of days since tool was used), as well as source species of vegetation, location of sourced parts within the plant (i.e., branch, stem) and height and diameter of stem or branch at point of material removal. Termite specimens from targeted mounds fixed in ethanol 80% were later identified by Rudolf Scheffrahn, University of Florida, USA. Samples from source plants were processed at camp with a plant drier for later identification by Frank Mbago, Botany Department, University of Dar es Salaam, Tanzania. RESULTS B group members have maintained their termite fishing tradition for more than four decades, as indicated by 2014–2016 records of 25 tools and 47 individual source plants, some of them sourced multiple times, comprising at least 10 species (Table 1). Source plants were found near 14 mounds (Figure 1 a–b), with 5 of these identified as Macrotermes michaelseni. One termite mound yielded tools on four dates (1 Dec 2014; 8 Dec 2014; 26 Nov 2016; 28 Nov 2016). Table 1. Plant species used by chimpanzees of B group at Mahale Mountains National Park to source termite fishing tools. 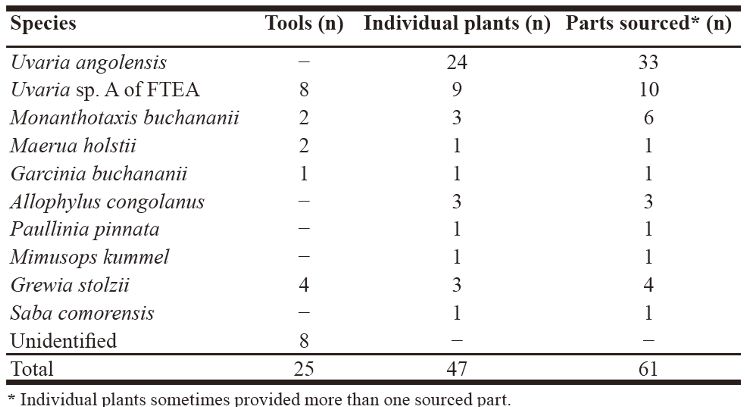

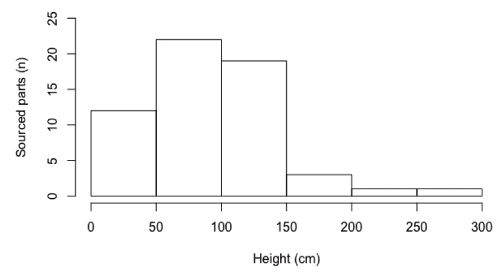
Recovered tools (n = 25) were traced back to 5 plant species, with 47% manufactured from an individual of Uvaria sp. A of FTEA, followed by Grewia stolzii (23.5%), with 3 tools manufactured from a single plant. Maerua holstii and Monanthotaxis buchananii were equally represented in the tool assemblage (11.7%), while Garcinia buchananii was the least represented (5.8%). All parts detached from plant sources (n = 61) were from bark extractions, with the majority sourced from Uvaria angolensis (54.0%) and Uvaria sp. A of FTEA (16.3%) (Table 1). All recovered tools were less than 24 h old, and made exclusively from bark (Figure 2 a–b). The mean length of tools was 38.5 cm (SD = 10.3, n = 25, range: 21.1–65.2), while mean diameter measured at medial point (approximately half-way along the tool) was 3.1 mm (SD = 1.0, n = 25, range: 1.5–5.1). Chimpanzees sourced the raw material (n = 58) mainly from branches (96.5%) and only rarely from stems of plants (3.4%). The vast majority (91.3%) was removed from up to 1.5 m above ground, at a mean height of 92.7 cm (SD = 50.8, n = 58, range: 5–280) (Figure 3). The mean diameter of the stem measured at the point from where the tool material was removed was 8.5 mm (SD = 2.1, n = 58, range: 15.5–5.3). 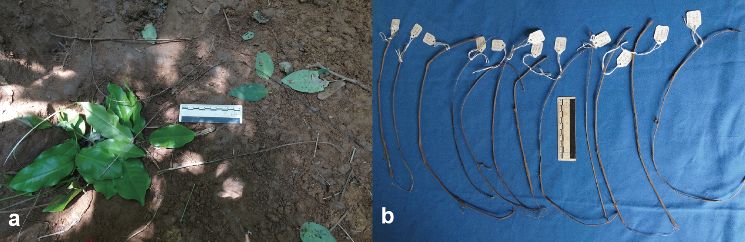
DISCUSSION This recent study shows that termite fishing has been maintained by the B group of Mahale chimpanzees for at least 42 years. While the physical characteristics of the tools, such as length and width, are similar with those reported decades ago (McGrew & Collins 1985), some differences emerged in relation to the species of termite prey and the type of material and species used for tool manufacture. While bark also was predominantly used in the past, other materials such as twig, sedge, vine, grass, bamboo, and leaf also were employed (Nishida & Uehara 1980; McGrew & Collins 1985). That only tools made from bark were recovered in this study may reflect a smaller sample size compared to previous tool assemblages (Nishida & Uehara 1980; McGrew & Collins 1985) (Table 2). Similarly, that only sources with evidence of bark extraction were found might be linked to the high fidelity that “scars” associated to these type of material extractions preserve on the source plant, compared to those associated with the detachment of other material such as twig, vine and grass, which hardly ever preserve (Pascual-Garrido, submitted). Table 2. Tools, plant source species and targeted prey associated with termite fishing by chimpanzees of B group at Mahale Mountains National Park. Bold = plant species sourced for raw material in more than one study. 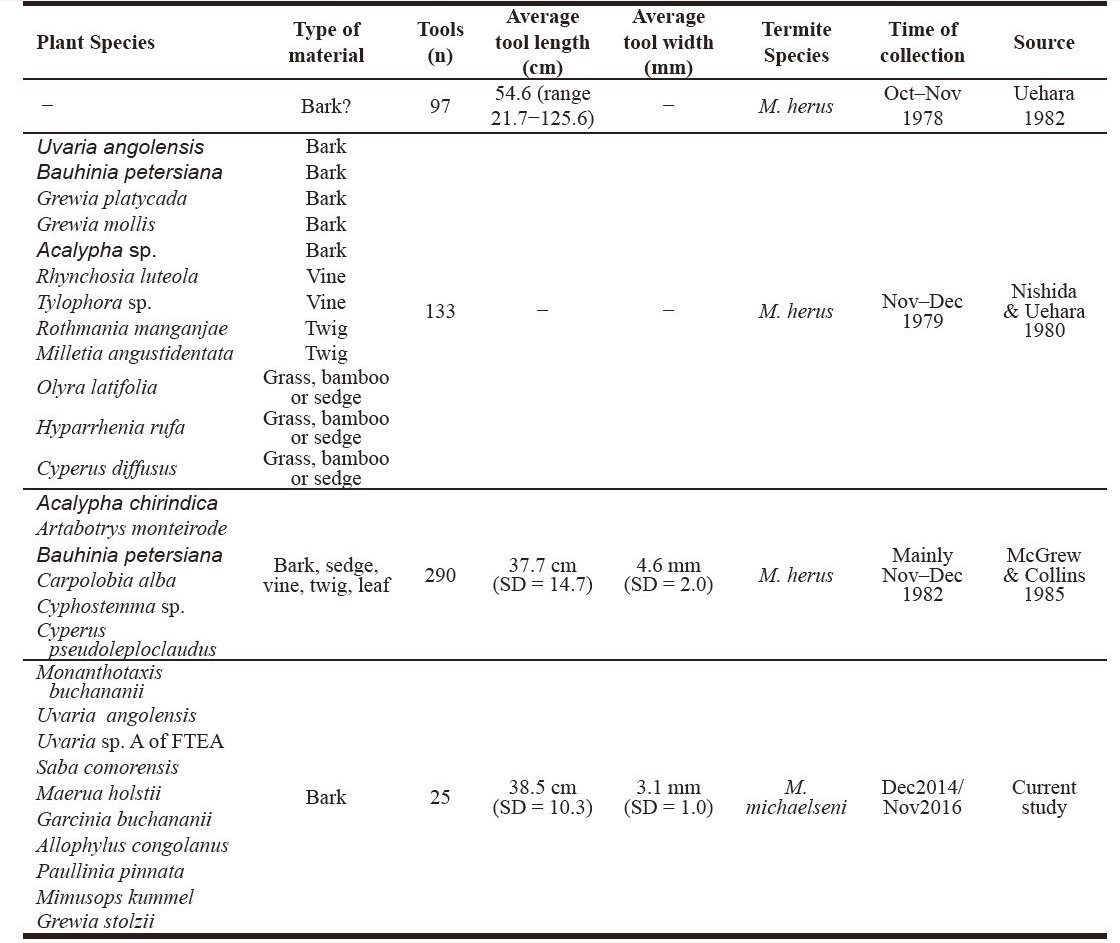
Alternatively, members of B group may have become exclusive bark users in recent decades. If so, then frequency differences in raw materials may reflect changes in their availability (McBeath & McGrew 1982). For example, the cessation of annual burning practice during the last decades might have allowed natural regeneration of woody vegetation near mounds (Collins & McGrew 1988). It also may be that historical variation in tool material preference is responsible (Hobaiter et al. 2014). Chimpanzees show material selectivity for manufacturing certain type of tools, including termite fishing probes (McBeath & McGrew 1982; Sanz & Morgan 2007; Almeida-Warren et al. 2017), a skill possibly transmitted across generations through social learning, such as via tool transfers (Sanz et al. 2014; Musgrave et al. 2016). However, without behavioral evidence of cultural transmission of tool material preference for this study community, this hypothesis remains untested. That measured length and width of the recovered tools seem to have been largely maintained for decades may be the product of socially learned skills (Sanz et al. 2014; Musgrave et al. 2016). Alternatively, this could simply be the result of ecological circumstances such as constancy of raw materials (McBeath & McGrew 1982). From the total of source species recorded in this study, only Uvaria angolensis has been used in the past, also as a source of bark (Nishida & Uehara 1980). However, Artabotrys monteiroae, Paulinnia pinnata and Grewia sp. were used by the formerly neighboring, but now extinct, K group to fish for Pseudoacanthotermes spiniger (Uehara 1982). Interestingly, Saba comorensis, Grewia sp, Monanthotaxis buchananii, Uvaria angolensis, and Uvaria sp. A of FTEA are currently sourced for termite fishing bark tools by Issa Valley chimpanzees living 85 km to the east (Almeida-Warren et al. 2017), while Uvaria angolensis, Allophylus congolanus and Saba comorensis constitute sources for termite fishing implements at Gombe, 200 km to the north (Pascual-Garrido, submitted). Different communities of chimpanzees using the same species of plants for tool material could result from their relative abundance. Alternatively, these species might have certain physical characteristics that make them ideal as tool raw material, i.e., optimal flexibility (Teleki 1974). This, in turn, may influence the likelihood that their usage is passed on via social transmission or by individual trial-and-error if the behavior occurs commonly enough. This could result in a possible exchange of members and their skills between communities (OMalley et al. 2012). However, given the fragmented landscape and presence of human population across the region, it is not clear that connectivity exists across ape populations (Piel et al. 2013). Chimpanzees sourced most raw material from branches close to the ground. However, some sources were 2.8 m high, indicating that chimpanzees sometimes climb up to reach desirable sources. Some sources even higher up may have not been visible to researchers. Values for mean height and diameter at which chimpanzees removed raw material at Bilenge are similar to those reported for chimpanzees at Issa Valley (Almeida-Warren et al. 2017). This might indicate that chimpanzees focus on where to source raw material, that is, certain heights or branch diameters from where bark easily can be detached (Sanz et al. 2004). Local traditions have been described for animal taxa as diverse as triggerfish, dolphins, whales and capuchin monkeys (for review see Fragaszy & Perry 2003). However, little is known about the exact time when traditions were invented, for how long they are maintained, or when they go extinct (Haslam et al. 2016). Thus, as McGrew (1992; p. 196) succinctly commented about the chimpanzees of Gombe: Termite fishing may just as well have been invented in 1959, the year before Jane Goodall arrived, or a million years ago. In any case, the current study contributes to the emerging paradigm of primate archaeology that aims to add time-depth to tool use behaviors of extant primate populations by identifying when and where tool use traditions are invented, modified or go extinct (Haslam et al. 2017). ACKNOWLEDGEMENTS Research permission for Mahale Mountains National Park was kindly granted by Tanzania National Parks (TANAPA), Tanzania Wildlife Research Institute (TAWIRI), and Tanzanian Commission for Science and Technology (COSTECH). The author is also thankful to: field assistance by Shaka and Singa Kabugonga; logistical support from Mahale Wildlife Conservation Society; plant identifications by Frank Mbago, Botany Department, University of Dar es Salaam; termite identification by Rudolf Scheffrahn, University of Florida, USA. Special thanks to Edward Kohi, Pius Kavana and Maria Mbogo for keeping my field equipment safe in Kigoma town; to Michio Nakamura for help with logistics in the field; to Volker Sommer for helpful comments on this manuscript; and to reviewer William McGrew for his constructive feedback. Financial support was provided by a Leverhulme Trust Early Career Fellowship and The Boise Trust Fund, University of Oxford. REFERENCES Almeida-Warren K, Sommer V, Piel AK, Pascual-Garrido A 2017. Raw material procurement for termite fishing tools by wild chimpanzees in the Issa valley, western Tanzania. Am J Phys Anthropol 164:292–304. https://doi.org/10.1002/ajpa.23269 Collins DA, McGrew WC 1988. Habitats of 3 groups of chimpanzees (Pan troglodytes) in western Tanzania compared. J Hum Evol 17:553–574. https://doi.org/10.1016/0047-2484(88)90084-X Fragaszy DM, Perry S (eds) 2003. The Biology of Traditions: Models and Evidence. Cambridge University Press, Cambridge, UK. Goodall J 1964. Tool using and aimed throwing in a community of free-living chimpanzees. Nature 201:1264–1266. https://doi.org/10.1038/2011264a0 Haslam M, Hernandez-Aguilar A, Arroyo A et al. 2017. Primate archaeology evolves. Nat Ecol Evol 1:1431–1437. https://doi.org/10.1038/s41559-017-0286-4 Haslam M, Luncz L, Pascual-Garrido A et al. 2016. Archaeological excavation of wild macaque stone tools. J Hum Evol 96:134–138. https://doi.org/10.1016/j.jhevol.2016.05.002 Hobaiter C, Poisot T, Zuberbühler K, Hoppitt W, Gruber T 2014. Social network analysis shows direct evidence for social transmission of tool use in wild chimpanzees. PLoS Biol 12:e1001960. https://doi.org/10.1371/journal.pbio.1001960 McBeath NM, McGrew WC 1982. Tools used by wild chimpanzees to obtain termites at Mt Assirik, Senegal: The influence of habitat. J Hum Evol 11:65–72. https://doi.org/10.1016/S0047-2484(82)80032-8 McGrew WC 1992. Chimpanzee Material Culture: Implications for Human Evolution. Cambridge University Press, Cambridge, UK. McGrew WC, Collins, DA 1985. Tool use by wild chimpanzees (Pan troglodytes) to obtain termites (Macrotermes herus) in the Mahale Mountains, Tanzania. Am J Primatol 9:47–62. https://doi.org/10.1002/ajp.1350090106 Musgrave S, Morgan D, Lonsdorf E, Mundry R, Sanz C 2016. Tool transfers are a form of teaching among chimpanzees. Sci Rep 6:34783. https://doi.org/10.1038/srep34783 Nakamura M, Corp N, Fujimoto M et al. 2013. Ranging behavior of Mahale chimpanzees: A 16 year study. Primates 54:171–182. https://doi.org/10.1007/s10329-012-0337-z Nishida T 1968. The social group of wild chimpanzees in the Mahali mountains. Primates 9:167–224. https://doi.org/10.1007/BF01730971 Nishida T, Uehara S 1980. Chimpanzees, tools and termites: Another example from Tanzania. Curr Anthropol 5:671–672. https://doi.org/10.1086/202545 OMalley RC, Wallauer W, Murray CM, Goodall J 2012. The appearance and spread of ant fishing among the Kasekela chimpanzees at Gombe. Curr Anthropol 53:650–663. https://doi.org/10.1086/666943 Pascual-Garrido A 2017. Termite fishing tools of wild chimpanzees and source plant scars: Towards an archaeology of the perishable. Manuscript submitted for publication. Pascual-Garrido A, Buba U, Nodza G, Sommer V 2012. Obtaining raw material: Plants as tool sources for Nigerian chimpanzees. Folia Primatol 83:24–44. https://doi.org/10.1159/000338898 Piel AK, Stewart FA, Pintea L et al. 2013. The Malagarasi river does not form an absolute barrier to chimpanzee movement in Western Tanzania. PLoS One 8:e58965. https://doi.org/10.1371/journal.pone.0058965 Sanz CM, Deblauwe I, Tagg N, Morgan DB 2014. Insect prey characteristics affecting regional variation in chimpanzee tool use. J Hum Evol 71:28–37. https://doi.org/10.1016/j.jhevol.2013.07.017 Sanz CM, Morgan DB 2007. Chimpanzee tool technology in the Goualougo Triangle, Republic of Congo. J Hum Evol 52:420–433. https://doi.org/10.1016/j.jhevol.2006.11.001 Sanz CM, Morgan DB, Gulick S 2004. New insights into chimpanzees, tools and termites from the Congo Basin. Am Nat 164:567–581. https://doi.org/10.1086/424803 Teleki G 1974. Chimpanzee subsistence technology: Materials and skills. J Hum Evol 3:575–594. https://doi.org/10.1016/0047-2484(74)90018-9 Uehara S 1982. Seasonal changes in the techniques employed by wild chimpanzees in the Mahale Mountains, Tanzania, to feed on termites (Pseudocanthotermes spiniger). Folia Primatol 37:44–76. https://doi.org/10.1159/000156020 Back to Contents |