|
<NOTE>
Savanna Chimpanzees (Pan troglodytes schweinfurthii) Consume and Share Blue Duiker (Philantomba monticola) Meat in the Issa Valley, Ugalla, Western Tanzania
Sebastian Ramirez-Amaya1, Edward McLester1,
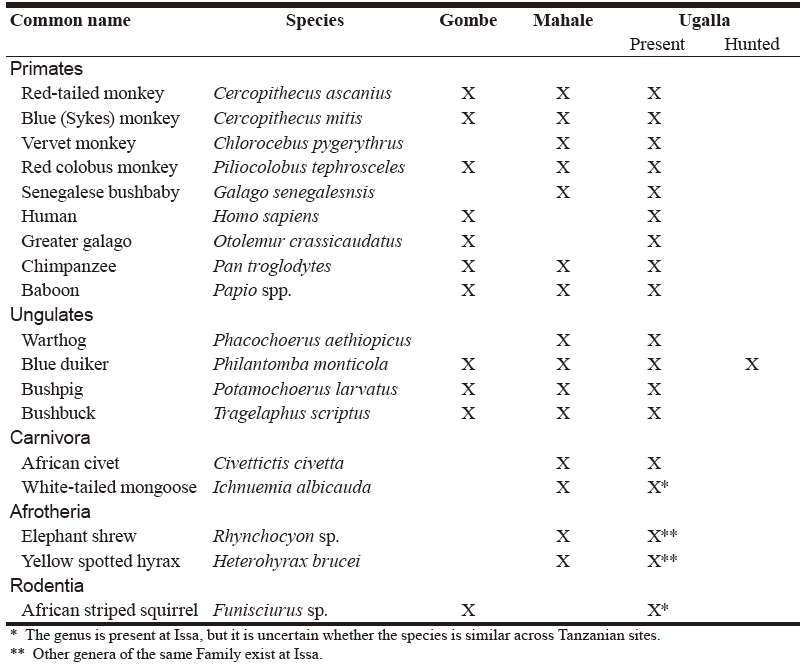
Fiona A. Stewart1,2, and Alex K. Piel1,3,* 1 Ugalla Primate Project 2 Department of Archaeology and Anthropology, University of Cambridge, UK 3 School of Natural Sciences and Psychology, Liverpool John Moores University, UK * Corresponding author INTRODUCTION Meat eating is pervasive across chimpanzee populations in Africa, with red colobus monkey (Piliocolobus spp.) being the most common prey (Boesch & Boesch 1989; Stanford et al. 1994a; Watts et al. 2012, Hosaka 2015) if sympatric in the same habitat. Besides colobus monkeys, chimpanzees consume a variety of other primates, including olive and yellow baboons (Papio spp.) and bushbabies (Galago spp.). In the forest habitats of western Tanzania chimpanzees have been reported to consume numerous different mammalian species: 18 at Mahale Mountains National Park (Uehara 1997; Hosaka 2015) and eight at Gombe National Park, whilst in the miombo woodland dominated Ugalla Region no direct observations have been recorded to date (Table 1). In West Africa, chimpanzees from Taï Forest, Ivory Coast consume eight different mammal species, all primates (Boesch & Boesch 1989). Wherever chimpanzees consume meat, it is almost always via hunting, as they rarely scavenge (Watts 2008).
Table 1. Species hunted at Gombe and Mahale, and whether they are present and hunted at Issa, modified from Goodall (1986) and Newton-Fisher (2014).
Habitat and wildlife diversity clearly influence potential prey for chimpanzees. In Fongoli, Senegal for example, chimpanzees live in a mosaic savanna landscape and prey on patas monkey (Erythrocebus patas) (Pruetz & Marshack 2009), a species that is rarely sympatric with otherwise mostly forest-dwelling chimpanzees. In Ugalla, two recent studies on the diet of wild chimpanzees each found only a paucity of animal tissue in over 1,200 combined samples. Yoshikawa and Ogawa (2015) reported a single case of bird bones and another of unidentified mammalian tissue in over 450 samples analyzed between 1995–2011 from the Nguye area, whilst Piel et al. (under revision) reported no mammalian tissue in over 800 samples collected from 2009–2014 from the Issa Valley. A reliance of faecal analysis to infer dietary behavior has well-discussed limitations (Boesch & Boesch 1989; McGrew et al. 2009; Phillips & McGrew 2013) and so direct observations are critical to revealing items that may be otherwise fully digested or rarely consumed. Where chimpanzees do capture prey, researchers have long been interested in meat-sharing (reviewed in Mitani & Watts 2001). Initial hypotheses described how males monopolized meat and used it as currency, trading it for mating opportunities either on a short or long-term basis, dubbed “meat for sex” (Stanford et al. 1994a, 1994b; Gomes & Boesch 2009). Others have argued that rather than sharing with females, meat-holders share preferentially instead with other males, using meat to build alliances with other males (Nishida et al. 1992; Mitani & Watts 2001). Finally, a third hypothesis suggested that individuals share meat to reduce the number of beggars, who are otherwise energetically expensive to continuously avoid (Gilby 2006). We report here on an opportunistic observation of chimpanzee consumption of blue duiker (Philantomba monticola) and subsequent acquisition of meat by party members in the Issa Valley, Ugalla, Tanzania. On 4 September, 2015 we observed multiple members of the Issa community feeding on the duiker carcass. We describe here this observation in the context of meat-eating of savanna chimpanzees and also the reliability of macro-analysis of faecal samples to infer dietary consumption. METHODS The Issa Valley is located in the Ugalla region, almost 100 km east of Lake Tanganyika (Figure 1) in western Tanzania. The study area extends over 85 km². The region is characterized by extreme seasonal variation: Annual rainfall averages 1,240 mm (range: 955–1,537) and the rainy season typically lasts from October to April, whilst the dry season (months with less than 100 mm of rainfall) lasts for five to six months, from April/May to September (Piel et al. 2015). The mosaic vegetation structure of the study area is dominated by miombo woodland (Brachystegia, Julbernardia, and Isoberlinia) interspersed with riverine forest, grasslands, and swamps. Russak (2014) recorded 42 mammal and 12 frugivorous bird species including lion (Panthera leo) and leopard (Panthera pardus). Most recently, researchers observed wild dog (Lycaon pictus) (McLester et al. unpublished data) in the area. 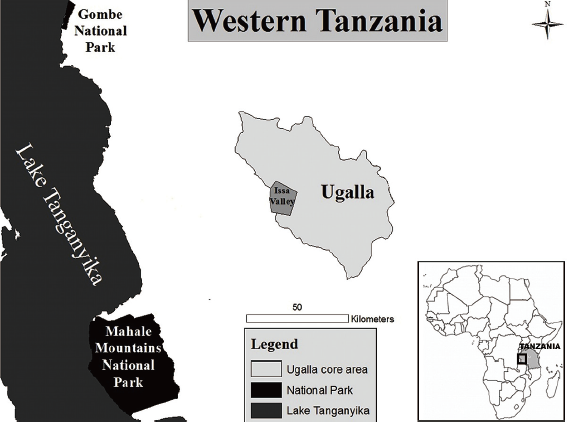
Figure 1. A map of western Tanzania, with the Issa study area as well as Gombe and Mahale Mountains National Parks identified (credit: L. Pintea, Jane Goodall Institute, USA). Since 2008, there has been a continuous research presence at Issa, and chimpanzees continue to show increased comfort with human presence, rarely fleeing approaching observers (Piel et al. unpublished data). All individuals are considered to be part of one community with an estimated home range of between 100–200 km² (Rudicell et al. 2011). As of September 2015, researchers had individually identified 14 different chimpanzees. To better understand chimpanzee dietary patterns, researchers collected faecal samples opportunistically and sluice samples in a near-by river. OBSERVATIONS At 0816 h EM and a field assistant encountered a party of at least five individuals who were foraging Julbernardia globliflora fruits in miombo woodland. Researchers followed the party into a riverine forest, with individuals periodically in and out of sight, when they heard a cacophony of chimpanzee vocalizations, including alarm barks. When researchers arrived, they encountered a party of nine chimpanzees and were able to approach within 10 m, although the chimpanzees were obscured in the canopy vegetation. Through a hole in the canopy, they identified an adult male climbing up whilst holding the carcass of a blue duiker in one of his hands (Figure 2). Six other individuals that clustered within 2 m around him followed him. At this time the carcass had already been dismembered, with only a portion remaining with the male in sight. The meat holder fed on and picked at the carcass whilst simultaneously allowing at least one other adult male as well as an adult female and her dependent offspring to feed from the meat by taking some pieces. Selectively, he chased and denied other individuals access to the meat. 
Figure 2. An adult male chimpanzee at Issa holds the blue duiker carcass (credit: E. McLester). After 10 min the meat holder left the tree, with some individuals in pursuit of him, whilst others remained in the tree. Researchers then saw the male climb an adjacent tree and disappear into the canopy, after which a chorus of pant hoots and screams were heard. There was a period of silence, and at 1015 h, researchers approached the location near the source of the vocalizations and identified three individuals consuming scraps of meat. By 1031 h, the chimpanzees had dispersed and researchers began searching for tissue remains and faecal samples in the surrounding area. None were recovered. The following day, researchers tracked a chimpanzee party for over 8 h. In that time they collected six faecal samples, three of which contained vertebrate remains: either hair or bone (tooth), or both (Figure 3). 
Figure 3. Mammalian (likely blue duiker) remains from chimpanzee faeces collected the day after the observed meat consumption (credit: S. Ramirez-Amaya). DISCUSSION Wild chimpanzees consume at least 40 different species of vertebrates across Africa (Newton-Fisher 2014) and numerous studies have addressed the role that meat might play as a nutritional resource (Boesch & Boesch-Achermann 2000), as a social tool to build alliances (Nishida et al. 1992), recruit mates (Stanford et al. 1994b) and to reciprocate meat sharing (Mitani & Watts 2001). For savanna-woodland chimpanzees in the Issa Valley our observations represent the first direct observation of mammalian consumption and expand the number of communities that are known to consume meat. During the last three decades of research on wild chimpanzees, discussion of chimpanzee predation focused on the consequences for arboreal prey, namely colobus populations (Stanford et al. 1994a; Hosaka et al. 2001; Mitani & Watts 2001; Newton-Fisher et al. 2002; Gomes & Boesch 2009). More recently, however, with the first habituation of savanna chimpanzees at Fongoli, more unorthodox sources of prey are being revealed, terrestrial and nocturnal, for example Erythrocebus (Pruetz & Marshack 2009) and Galago (Pruetz & Bertolani 2007). Now at Issa, we report an observation that also suggests a terrestrial capture. Issa chimpanzees are not the only community to consume antelopes. The chimpanzees of Mahale Mountains also consume blue duikers (Takahata et al. 1984). What remains poorly understood is what drives prey selection and hunting frequency. 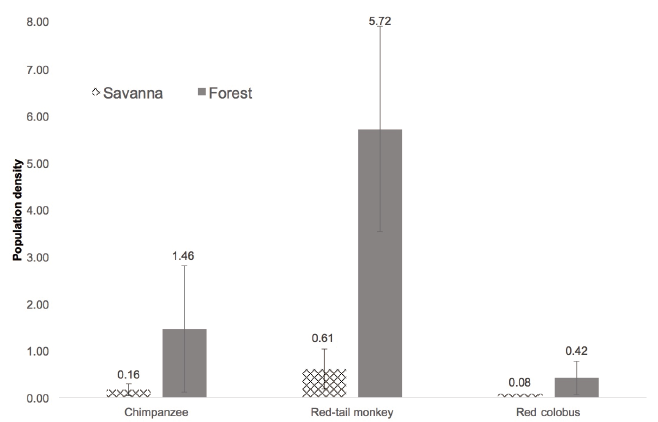
Figure 4. Population density variation between forest-dwelling and savanna primates, with error bars indicating standard deviations (source: Plumptre & Reynolds 1996; Chapman & Lambert 2000; Moyer et al. 2006; Rode et al. 2006; Davenport et al. 2007; Petre et al. 2007; Kouakou et al. 2009; Moore & Vigilant 2013; Samson & Hunt 2014; Piel et al. 2015). A single record exists from Issas red colobus population, hence the lack of error bars. There are at least three potential explanations for prey selection and hunting frequency variability. First, one reason for lower prevalence of meat-eating at Issa may be the population density of both chimpanzees and potential prey (Figure 4). For example, chimpanzees and red-tailed monkeys live at very low densities, reducing the likelihood of encounters between the species. It may be that with less forest available, and subsequently lower monkey density (Figure 4), chimpanzees living in more open habitats exploit alternative prey sources. More data on prey availability and preference might resolve whether an environmental explanation is sufficient. Second, an alternative explanation in terms of culture might be explored: According to Boesch & Boesch (1989), Taï forest-chimpanzees never eat blue duikers even if they capture them, although blue duikers are fairly common in their habitat, whereas Issa chimpanzees have not been reported to capture or eat arboreal prey, despite the fact that chimpanzees and arboreal monkeys live sympatric at Issa. Third, macro-analysis may be insufficient for detecting animal tissue. Whilst it reveals much about chimpanzee diet (Phillips & McGrew 2013), Boesch & Boesch (1989) have outlined its limitations. That no mammalian tissue was observed in an analysis of over 1200 faecal samples across Ugalla suggests that either meat-eating is an extremely rare event, or else not all items that chimpanzees consume are detectable in faeces. We suspect it is the former, and as chimpanzee habituation improves in the coming months, we anticipate observing more chimpanzee predation events. ACKNOWLEDGEMENTS The authors thank the Tanzanian Wildlife Research Institute (TAWIRI) and the Commission for Science and Technology (COSTECH) for permission to conduct research in the Issa Valley, and field assistants at the Ugalla Primate Project for efforts to track chimpanzees. Support for UPP comes from the UCSD/Salk Center for Academic Research and Training in Anthropogeny (CARTA). REFERENCES Boesch C, Boesch H 1989. Hunting behavior of wild chimpanzees in the Taï National Park. Am J Phys Anthropol 78: 547–573. Boesch C, Boesch-Achermann H 2000. The Chimpanzees of the Taï Forest. Oxford University Press, Oxford. Chapman CA, Lambert JE 2000. Habitat alteration and the conservation of African primates: Case study of Kibale National Park, Uganda. Am J Primatol 50: 169–185. Davenport TRB, Mpunga NE, Machaga SJ 2007. Census and conservation assessment of the red colobus newly-discovered, threatened and extinct populations. Primate Conserv 22: 97–105. Gilby IC 2006. Meat sharing among the Gombe chimpanzees: Harassment and reciprocal exchange. Anim Behav 71: 953–963. Gomes CM, Boesch C 2009. Wild chimpanzees exchange meat for sex on a long-term basis. PloS One 4: e5116. Goodall J 1986. The Chimpanzees of Gombe: Patterns of Behaviour. Harvard University Press, Cambridge, MA. Hosaka K 2015. Hunting and food sharing. In: Mahale Chimpanzees: 50 Years of Research. Nakamura M, Hosaka K, Itoh N, Zamma K (eds), Cambridge University Press, Cambridge, pp. 274–290. Hosaka K, Nishida T, Hamai M, Matsumoto-Oda A, Uehara S 2001. Predation of mammals by the chimpanzees of the Mahale Mountains, Tanzania. In: All Apes Great and Small, Volume I: African Apes. Galdikas B, Nancy Briggs E, Sheeran L, Shapiro G, Goodall J (eds), Kluwer, New York, pp. 107–130. Kouakou CY, Boesch C, Kuehl K 2009. Estimating chimpanzee population size with nest counts: Validating methods in Taï National Park. Am J Primatol 457: 447–457. McGrew WC, Marchant LF, Phillips CA 2009. Standardised protocol for primate faecal analysis. Primates 50: 363–366. Mitani JC, Watts DP 2001. Why do chimpanzees hunt and share meat? Anim Behav 61: 915–924. Moore DL, Vigilant L 2013. A population estimate of chimpanzees (Pan troglodytes schweinfurthii) in the Ugalla region using standard and spatially explicit genetic capture-recapture methods. Am J Primatol 76: 335–346. Moyer D, Plumptre AJ, Pintea L, Hernandez-Aguilar A, Moore J, Stewart F, Davenport TRB, Piel A, Kamenya S, Mugabe H, Mpunga N, Mangoka N 2006. Surveys of Chimpanzees and Other Biodiversity in Western Tanzania. Report submitted to USF&W, Great Apes Fund. Newton-Fisher NE 2014. The hunting behavior and carnivory of wild chimpanzees. In: Handbook of Paleoanthropology. Springer, Berlin Heidelberg, pp. 1661–1691. Newton-Fisher NE, Notman H, Reynolds V 2002. Hunting of mammalian prey by Budongo forest chimpanzees. Folia Primatol 73: 281–283. Nishida T, Hasegwa T, Hayaki H, Takahata Y, Uehara S 1992. Meat-sharing as a coalition strategy by an alpha male chimpanzee? In: Topics in Primatology, Vol1. Human Origins. Nishida T, McGrew WC, Marler P, Pickford M, de Waal F (eds), Tokyo University Press, Tokyo, pp. 159–174. Petre CA, Huynen MC, Beudels-Jamar R 2007. First assessment of chimpanzee (Pan troglodytes troglodytes) density and bedding behaviour in the Pongara National Park, Gabon. Pan Afr News 14: 22–27. Phillips CA, McGrew WC 2013. Identifying species in chimpanzee (Pan troglodytes) feces: A methodological lost cause? Int J Primatol 34: 792–807. Piel AK, Lenoel A, Johnson C, Stewart FA 2015. Deterring poaching in western Tanzania: The presence of wildlife researchers. Global Ecol Conserv 3: 188–199. Plumptre AJ, Reynolds V 1996. Censusing chimpanzees in the Budongo Forest, Uganda. Int J Primatol 17: 85–99. Pruetz JD, Bertolani P 2007. Savanna chimpanzees, Pan troglodytes verus, hunt with tools. Curr Biol 17: 412–417. Pruetz JD, Marshack JL 2009. Savanna chimpanzees (Pan troglodytes verus) prey on patas monkeys (Erythrocebus patas) at Fongoli, Senegal. Pan Afr News 16: 15–17. Rode KD, Chapman CA, Mcdowell LR, Stickler C 2006. Nutritional correlates of population density across habitats and logging intensities in redtail monkeys (Cercopithecus ascanius). Biotropica 38: 625–634. Rudicell RS, Piel AK, Stewart F, Moore DL, Learn GH, Li Y, Takehisa J, Pintea L, Shaw GM, Moore J, Sharp PM, Hahn BH 2011. High prevalence of simian immunodeficiency virus infection in a community of savanna chimpanzees. J Virol 85: 9918–9928. Russak SM 2014. Using patch focals to study unhabituated dry-habitat chimpanzees (Pan troglodytes schweinfurthii) and sympatric fauna at Issa, Ugalla, Tanzania. Int J Primatol 35: 1202–1221. Samson DR, Hunt KD 2014. Is chimpanzee (Pan troglodytes schweinfurthii) low population density linked with low levels of aggression? Pan Afr News 21: 15–17. Stanford CB, Wallis J, Matama H, Goodall J 1994a. Patterns of predation by chimpanzees on red colobus monkeys in Gombe National Park, 1982–1991. Am J Phys Anthropol 94: 213–228. Stanford CB, Wallis J, Mpongo E, Goodall J 1994b. Hunting decisions in wild chimpanzees. Behaviour 131: 1–18. Takahata Y, Hasegwa T, Nishida T 1984. Chimpanzee predation in the Mahale Mountains from August 1979 to May 1982. Int J Primatol 5: 212–233. Uehara S 1997. Predation on mammals by the chimpanzee (Pan troglodytes). Primates 38: 193–214. Watts DP 2008. Scavenging by chimpanzees at Ngogo and the relevance of chimpanzee scavenging to early hominin behavioral ecology. J Hum Evol 54: 125–133. Watts DP, Potts KB, Lwanga JS, Mitani JC 2012. Diet of chimpanzees (Pan troglodytes schweinfurthii) at Ngogo, Kibale National Park, Uganda, 2: Temporal variation and fallback foods. Am J Primatol 144: 130–144. Yoshikawa M, Ogawa H 2015. Diet of savanna chimpanzees in the Ugalla area, Tanzania. Afr Stud Monogr 36: 189–209. Back to Contents |