|
<NOTE>
Association of a Young Emigrant Female Bonobo during an Encounter with her Natal Group
Kazuya Toda, Tetsuya Sakamaki, Nahoko Tokuyama & Takeshi Furuichi
Primate Research Institute of Kyoto University, Japan INTRODUCTION Sex-biased dispersal is an almost ubiquitous feature of the life histories of group-living mammals (Handley & Perrin 2007). Most primates resemble other mammals in showing male-biased dispersal, which is considered to be related to the fact that female reproductive success is limited primarily by nutritional constraints in mammals where females bear the major burden of investment of their offspring, such as birth and nursing (Greenwood 1980). On the other hand, societies of chimpanzees and bonobos show clear female-biased dispersal: females usually transfer between groups upon reaching sexual maturity (Thompson 2013; Sakamaki et al. in press). Although inbreeding avoidance may be a plausible ultimate factor of sex-biased dispersal in mammals (Pusey 1980), it has not been well understood why females emigrate from natal groups in these species. Similarly, proximate factors for female transfer remain poorly understood due to the difficulty of successive observations of young females before and after their transfer. Therefore, more information is needed on the process of female transfer between groups. Bonobo societies maintain relatively less antagonistic relations between groups, and affiliative interactions are observed between members of different groups during encounters (Idani 1990; Furuichi 2011). Young females often migrate between groups during inter-group encounter not only once but also repeatedly (Sakamaki et al. in press). Association patterns of young females during the encounters should reflect their motivation to emigrate into another group or to remain in the current group. Here, we report observations of a young emigrant female during an encounter between her current group and her natal group over four successive days. We show the association patterns and social interactions of the young female with members of both groups. MATERIALS & METHO DS Observations for this study were made at Wamba in the northern sector of the Luo Scientific Reserve in DR Congo (Furuichi et al. 1998). Studies at Wamba started in 1973, but were interrupted from 1996 to 2002 due to the civil war. Research was resumed in 2003, and one study group (E1) has been followed on a daily basis since. Researchers had also observed a western adjacent group (P) until 1996, and observation of this group resumed with intensive habituation on a daily basis in September 2010. This group was renamed P-east (Pe) to differentiate it from another western group, P-west (Pw). All animals in Pe group were identified by August 2011. The E1 group consisted of 36 animals and the Pe group consisted of 26 animals in August 2014. A young female (Puffy) was first identified in the Pe group in November 2010 (Figure 1). From our observations, we assumed that an adult female (Pao) was her mother and a female infant (Pipi) was her younger sister. She was estimated to be 7 years old from her body size and the inter-birth interval with Pipi, whose age was estimated at 2 years (the inter-birth interval in bonobos is considered to be approximately 4.8 years; Furuichi et al. 1998). She emigrated from the Pe group to E1 group on October 12, 2013 during an encounter between the groups, and has remained with the E1 group ever since. The next encounter between the Pe and E1 groups occurred on August 16, 2014, when Puffy met members of her natal group for the first time since her emigration. 
Figure 1. Puffy in March 2015. On August 16, 2014, an inter-group encounter occurred in the area that Pe group usually ranged. Puffy arrived at the area with E1 members. The groups were repeatedly within sight of each other, and when separated were usually within earshot until August 19. One of the authors (KT) recorded the behaviors of Puffy via focal animal sampling for approximately 3 hours per day during these four days. KT also recorded the identities of animals that were observed within 5 m using scan sampling at 5 minute intervals during focal animal observations. A total of 144 scan samples were recorded over four days, and we used data from 116 scan samples excluding 28 scan samples when KT lost sight of Puffy from the analysis (25 samples on August 16, 31 samples on August 17, 27 samples on August 18, 33 samples on August 19). Frequencies of social grooming with Puffy were determined from these scan data. We used the records of animals within 5 m of Puffy to compare the mean number of animals in all scans (called per-scan number hereafter) and the actual number of animals on each day (called all-day number hereafter) between the E1 group (27 animals) and Pe group (23 animals), excluding infants (<4 years old). The all-day number was calculated by counting animals found in the proximity of Puffy at least once in a day. RESULT S & DISCUSSION Figure 2 compares the mean number of E1 animals and Pe animals found in Puffys proximity in 116 scans. In these scans, an average of 0.11 animals from the E1 group and 0.29 animals from the Pe group were found within 1 m of Puffy, while 0.70 animals from the E1 group and 0.53 animals from the Pe group were recorded within 5 m. A statistical test where we resampled one scan for 30 min to avoid auto-correlation did not show a significant difference in the per-scan number between the E1 and Pe members at either distance. Figure 3 compares the all-day number of animals found in the proximity of Puffy on each day between the E1 and Pe groups. There was no difference between the numbers of E1 and Pe members within 1 m of Puffy. However, while there were no specific animals among E1 members regularly found within 1 m, two members of the Pe group were very frequently found within 1 m of Puffy (Marie, a young adult female: 24 times, 71% of all scan records of Pe animals in 1 m proximity; and Michio, a juvenile male: 6 times, 18% of all scan records). Puffy exchanged social grooming with these two animals more frequently (11 scans with Marie and 3 scans with Michio out of 16 scans with social grooming) than with E1 members. On the other hand, 24 animals from the E1 group (89% of all E1 members) and 11 animals from the Pe group (48% of all Pe members) were found within 5 m at least once in all scans (116 scans). These results suggest that Puffy ranged more with E1 members than Pe members during the focal observation. Thus, Puffy appeared to associate with E1 members rather than Pe members for the four days of the study period, while she had much more frequent grooming interactions with specific animals of her natal group. 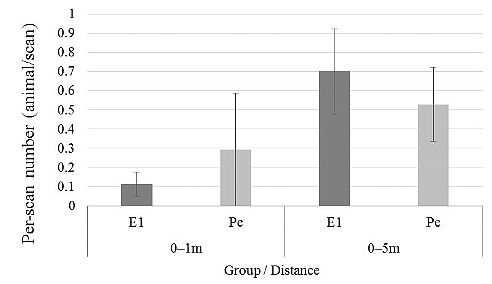
Figure 2. Comparisons of per-scan number of animals found in Puffys proximity (in 116 scans) between the E1 and Pe groups. This figure shows how frequently E1 or Pe animals were observed within 1 m or 5 m of Puffy. Error bars show the standard deviation. We resampled one scan for 30 min (N = 24; 144 scans/6 scans) to avoid auto-correlation and performed a Wilcoxon signed rank test to examine the difference between the number of E1 animals and that of Pe animals within 1 and 5 m. There were no significant differences between the number of E1 and Pe members either within 1 m (T = 31, N = 24, n.s.) or within 5 m (T = 70, N = 24, n.s.). 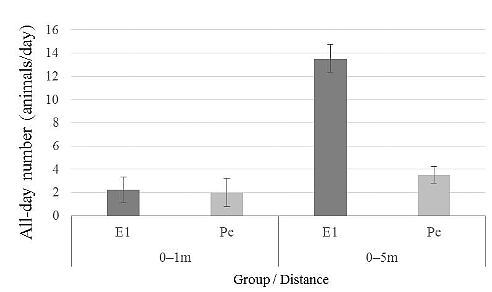
Figure 3. Comparisons of the all-day number of animals found in Puffys proximity for each day of observation (N = 4 days). This figure shows how many E1 or Pe animals were recorded within 1 m or 5 m of Puffy on each day. Error bars show the standard deviation. Although both Pao and Puffy were observed on all four days, Pao was observed within 5 m of Puffy in only one of the 116 scans conducted. Puffy was observed interact with Pao on the first day of the group encounter: NT observed that Puffy played with Pao and begged her for food during ad libitum observation. However, no social interactions between them were observed in focal observations. These results suggest that neither Puffy nor Pao paid particularly attention to one another during the observation. Puffy was frequently observed to associate with Pao before her emigration in 2013, although she rarely groomed with her. Therefore, the relationship between Puffy and Pao appeared to have changed gradually before her emigration. This was the first observation of an emigrant female during an encounter with her natal group members at Wamba. Interestingly, Puffy seemed estranged from her mother and members of her natal group after 10 months of separation although she interacted with some young animals of her natal group. She may have already become the member of the E1 group in the 10 months. This observation suggests that Puffy has no motivation to return to her natal group although it cannot be asserted that the cause is a change of her group identity into the new group. Habituation of several other groups adjacent to E1 and Pe is currently proceeding at Wamba. We expect that the accumulation of similar observations will contribute to an increased understanding of how female bonobos decide on transfers between groups. ACKNOWLEDGEMENTS This research was financially supported by the Leading Graduate program in Primatology and Wildlife Science of Kyoto University (to KT), the Japan Society for the Promotion of Science (JSPS) Asia-Africa Platforms (to TF), the Japan Ministry of the Global Environment Research Fund (to TF), JSPS Grants-in-Aid for Strategic Young Researcher Overseas Visits Program for Accelerating Brain Circulation (to Primate Research Institute, Kyoto University), and JSPS HOPE project of the Primate Research Institute of Kyoto University, JSPS Research Fellowships for Young Scientist (to NT). We thank the DR Congo Ministry of Science for permission to conduct research at Wamba. We also thank the local staff and others at Wamba for their support during our study. REFERENCES Furuichi T 2011. Female contributions to the peaceful nature of bonobo society. Evol Anthropol 20:123142. Furuichi T, Idani G, Ihobe H, Kuroda S, Kitamura K, Mori A, Enomoto T, Okayasu N, Hashimoto C, Kano T 1998. Population dynamics of wild bonobos (Pan panisucus) at Wamba. Int J Primatol 19:10291043. Greenwood PJ 1980. Mating systems, philopatry and dispersal in birds and mammals. Anim Behav 28:11401162. Handley L, Perrin N 2007. Advances in our understanding of mammalians sex-biased dispersal. Molecul Ecol 16:15591578. Idani G 1990. Relations between unit-groups of bonobos at Wamba, Zaire: Encounters and temporary fusions. Afr Stud Monogr 11:153186. Pusey AE 1980. Inbreeding avoidance in chimpanzees. Anim Behav 28:543552. Sakamaki T, Behncke I, Laporte M, Mulavwa M, Ryu H, Takemoto H, Tokuyama N, Yamamoto S, Furuichi T in press. Intergroup transfer of females and social relationships between immigrants and residents in bonobo (Pan paniscus) societies. In: Dispersing Primate Females: Life History and Social Strategies in Male-Philopatric Species. Furuichi T, Yamagiwa J, Aureli F (eds), Springer, Tokyo. Thompson ME 2013. Reproductive ecology of female chimpanzees. Am J Primatol 75:222237. Back to Contents |