|
<NOTE>
Responses of Wild Chimpanzees to Fresh Carcasses of Aardvark (Orycteropus afer) in Mahale
Kazuhiko Hosaka1, Eiji Inoue2 & Mariko Fujimoto3
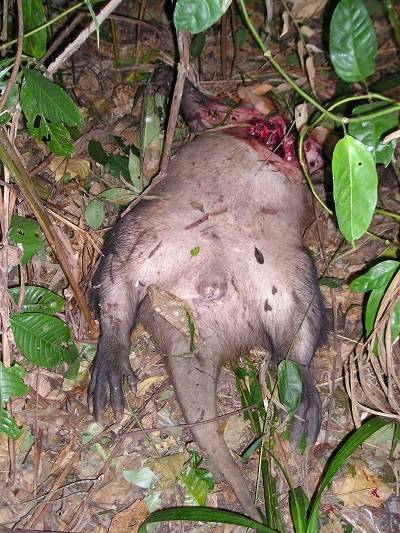
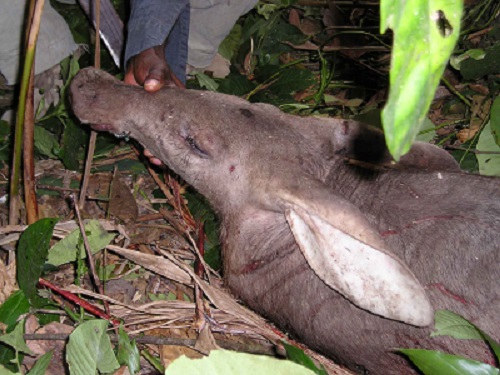
1 Kamakura Women's University, Japan 2 Department of Zoology, Graduate School of Science, Kyoto University, Japan 3 Graduate School of Asian and African Area Studies, Kyoto University, Japan INTRODUCTION Despite more than 50 years of research on wild chimpanzees, there has been no record of encounters between them and aardvarks (Orycteropus afer). The habitats of aardvarks are distributed widely in sub-Saharan Africa and cover most of the narrower habitats of chimpanzees (Lindsey et al. 2008). However, aardvarks are typically nocturnal, foraging for termites, ants, and larvae at night, while chimpanzees are typically diurnal, hence the lack of recorded encounters between the two species. This study reports the first two recorded observations of the rare event of chimpanzees encountering aardvark carcasses, which occurred within 1 month. This provides valuable insight on two topics. The first topic is the reaction of chimpanzees to nonconspecific carcasses. The observations provide a variety of clues about chimpanzees' emotional traits and their cognition of potential dangers such as predation and illness. Several studies (Teleki 1973; Boesch 1991; Hosaka et al. 2000) reported that chimpanzees showed intense emotions of fear and confusion toward dead bodies of mature members belonging to the same unit-group. Early, harsh “wraa” calls and late, feeble “huu” calls (Goodall 1986) seem to be typically expressed during such encounters. The same calls are emitted when chimpanzees see or hear large carnivores such as leopards (Panthera pardus) and lions (Panthera leo) (Hiraiwa-Hasegawa et al. 1986; Tsukahara 1993). The second topic is the scavenging tendency of chimpanzees. Watts (2008) reviewed published cases of scavenging by wild chimpanzees and argued that chimpanzees avoid scavenging because fresh carcasses are rare in the forest and chimpanzees may not recognize certain species as potential prey, whereas Mahale chimpanzees sporadically scavenged fresh carcasses of their prey species, such as bushbuck (Tragelaphus scriptus), killed and cached by leopards (Hasegawa et al. 1983; Hosaka et al. 2001). In contrast, several reports (e.g. Boesch & Boesch 1989; Hirata et al. 2001) suggest that the western chimpanzees do not eat mammals absent from their prey species list at the respective study sites; they did not consume a blue duiker (Cephalophus monticola) or a western tree hyrax (Dendrohyrax dorsalis) accidentally captured and killed, but only toyed with them. METHODS Observations of wild chimpanzees (M-group) were made in the Mahale Mountains National Park, Tanzania, on August 17 and September 4, 2005. Audiovisual data from video tapes and field notes taken by the authors were analyzed to obtain a sequence of events. Sympatric aardvarks have never been studied in detail but their nocturnal activities were observed in some camera-trap studies (Moyer 2006; Nakamura, personal communication). OBSERVATIONS 1. August 17, 2005. At approximately 0930 h, just before moving to forage, a group of chimpanzees encountered a dead aardvark that appeared to be a subadult female (> 30 kg) in the leafy bush near the river junction of the Manya and the Kasoje. Initially, several individuals, including OR (adolescent male) and SY (adult female), emitted “wraa” calls so harshly that others going eastward turned back westward to reach the site. All 21 individuals in the foraging group gathered at the site. Although there was a diversity of responses among individual chimpanzees, there were some tendencies observed for age-sex classes (Table 1). Adolescent males showed the highest intensity and ambiguity in their responses. OR, who emitted “wraa” calls most often, would not approach the carcass and remained still on a tree approximately 20 m distant. Three other adolescent males, PR, CD, and MC, boldly approached the carcass, and inspected it thoroughly, sometimes touching it. Similar explorative behaviors were rare among other age-sex individuals, although three adult/adolescent females touched the carcass. Other individuals, including four adult males, approached to watch the carcass and stayed for a while without emotional confusion. At approximately 1030 h, their emotional reactions gradually subsided, with a sporadic production of “huu” calls. At 1045 h, the last remaining individual, GW (old female), left the site. Evidently, not a single individual attempted to taste the carcass meat. The aardvark was presumed to have been freshly killed by a leopard, a couple of hours before being encountered. There were many fresh claw wounds on the surface of the body (Figure 1). A large fatal open wound was observed on the chest and some meat had been eaten by the killer. Fresh blood remained wet. Only a few flies had reached the body. Later in the afternoon, one of the authors (E.I.) went back to the site to find that the carcass had disappeared. We presume that the leopard had returned to the carcass after the chimpanzees and researchers had left the site, and removed it. According to Bailey (1990), a leopard will not move far from a cached carcass, often concealing itself and resting in the vicinity, so as to return to feed when its appetite is renewed.
2. September 4, 2005. At approximately 1300 h, in arid bush near the sunexposed, dry bed of the Ikola River, chimpanzees encountered another aadvark carcass; this one was foul-smelling and putrid. There was no indication as to the cause of death. It appeared to have been dead for 4 or 5 days. No “wraa” calls were heard (Table 1). Several individuals emitted “huu” calls as they walked in single file toward the carcass, but most of them passed the carcass without close inspection. Only three of 26 individuals— CD, OS (juvenile male), and LZ (juvenile female) —made a detour to inspect the carcass. Only CD touched the carcass. The two juveniles (OS and LZ) showed intense, ambiguous emotions, reflecting curiosity and fear toward the carcass, repetitively approaching and retreating. Threatening behaviors toward the carcass were observed in this case only. The chimpanzees sometimes approached the carcass to inspect it and attempted to repel flies by shaking their hands (Table 1). The two juveniles would rush away each time they threatened the carcass. Neither of them attempted to bite the carcass or eat any meat. 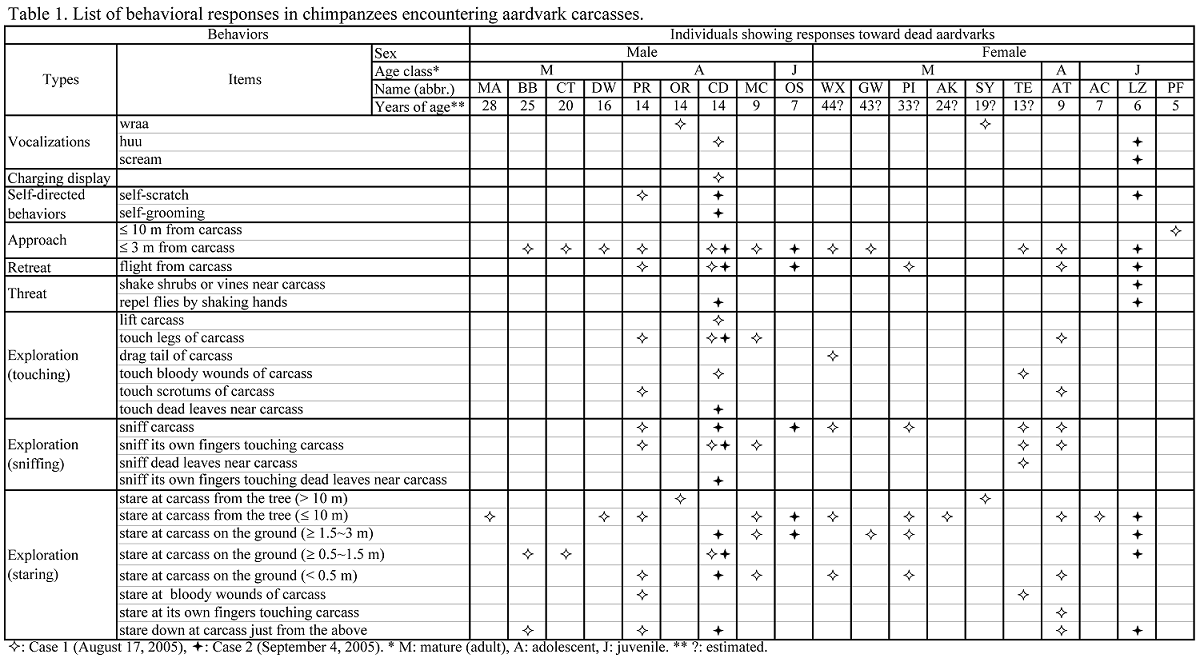
DISCUSSION As Hosaka et al. (2000) have suggested, there is no substantial difference in chimpanzee responses when they encounter conspecific or non-conspecific dead bodies. They show emotions by emitting “wraa” calls or “huu” calls. They approach, watch, sniff, and touch the carcass for inspection, while they are ready to run away from the carcass with a slight stimulus or none. It seems plausible that these behaviors reflect “general curiosity” in wild chimpanzees. Unusual or unfamiliar objects often motivate an animal's explorative behavior, since it may be adaptive in learning of risks and dangers. Fear arises at the same time and motivates the animal's retreat because the explored objects may be risky or dangerous to it. However, two differences may be noticed in chimpanzees' responses when they encounter the carcasses of either familiar conspecific or non-conspecific mammals. First, chimpanzees did not show as much curiosity about the putrid non-conspecific carcass (this study) as they showed about a putrid conspecific carcass (Hosaka et al. 2000). The identity of the carcass and the long-term relationships with the dead individual may have affected their responses to a certain degree. Second, chimpanzees showed more curiosity about the fresh non-conspecific carcass (this study) than they showed about the fresh carcass of a conspecific animal that died from a respiratory disease (Hosaka et al. 2000). Cause of death may have motivated different responses. If chimpanzees find a carcass freshly killed by a leopard, this may motivate them to be alert and to explore it for more information about the predator. Finally, this study provided valuable information as to the stenophagous nature of chimpanzee meat-eating habits. Ihobe (1993) insisted that chimpanzees are stenophagous meat-eaters in contrast to humans, who have evolved as euryphagous meat-eaters after acquiring a “prey image” for any type of mammal meat and beginning to rely on a “scavenging life” in savannah (Speth 1989). It is likely that chimpanzees did not eat the fresh aardvark carcass simply because they had no prey image for this nocturnal mammal that they would not normally encounter during their ranging in the daytime. It seems easier for chimpanzees to develop a prey image for diurnal mammals (e.g. arboreal monkeys), with which they have a high encounter rate (Uehara 1997; Hosaka et al. 2001). ACKNOWLEDGEMENTS The fieldwork for this study was supported by a JSPS Grant-in-Aid for Scientific Research (A) (1) (#16255007 to Toshisada Nishida). Analysis of the data from videos and field notes was supported by a MEXT Grant-in-Aid for Young Scientists (B) (#15770159 to Kazuhiko Hosaka). We are indebted to the Tanzania Commission for Science and Technology (COSTECH), the Tanzania Wildlife Research Institute (TAWIRI), and the Tanzania National Parks (TANAPA) for permission to undertake research on chimpanzees in the Mahale Mountains National Park. We also thank all the local assistants for their cooperation in the daily tracking of chimpanzees. REFERENCES Bailey TN 1990. The African Leopard. Columbia University Press, New York. Boesch C 1991. The effect of leopard predation on grouping patterns in forest chimpanzees. Behaviour 117:220–242. Boesch C, Boesch H 1989. Hunting behavior of wild chimpanzees in the Taï National Park. Am J Phys Anthropol 78:547–573. Goodall J 1986. The Chimpanzees of Gombe: Patterns of Behavior. Belknap, Cambridge, MA. Hasegawa T, Hiraiwa M, Nishida T, Takasaki H 1983. New evidence on scavenging behavior in wild chimpanzees. Curr Anthropol 24:231–232. Hiraiwa-Hasegawa M, Byrne RW, Takasaki H, Byrne JM 1986. Aggression toward large carnivores by wild chimpanzees of Mahale Mountains National Park, Tanzania. Folia Primatol 47:8–13. Hirata S, Yamakoshi G, Fujita S, Ohashi G, Matsuzawa M 2001. Capturing and toying with hyraxes (Dendrohyrax dorsalis) by wild chimpanzees (Pan troglodytes) at Bossou, Guinea. Am J Primatol 53:93–97. Hosaka K, Matsumoto-Oda A, Huffman MA, Kawanaka K 2000. Reactions to dead bodies of conspecifics by wild chimpanzees in the Mahale Mountains, Tanzania. Primate Res 16:1–15, in Japanese with English summary. Hosaka K, Nishida T, Hamai M, Matsumoto-Oda A, Uehara S 2001. Predation of mammals by the chimpanzees of the Mahale Mountains, Tanzania. In: All Apes Great and Small, vol. 1: African Apes. Galdikas BMF, Nancy Briggs E, Sheeran L, Shapiro G, Goodall J (eds), Kluwer Academic, New York, pp. 107–130. Ihobe H 1993. Meat and beast: Prey images of bononos, chimpanzees, and humans. J Afr Stud 42:61–68, in Japanese.
Lindsey P, Cilliers S, Griffin M, Taylor A, Lehmann T,
Rathbun G (IUCN SSC Afrotheria Specialist Group)
2008. Orycteropus afer. The IUCN Red List of Threatened
Species. Version 2014.3. Moyer DC 2006. Biodiversity of Mahale Mountains National Park, Tanzania. Wildlife Conservation Society, New York. Speth JD 1989. Early hominid hunting and scavenging: the role of meat as an energy source. J Hum Evol 18:329–343. Teleki G 1973. Group response to the accidental death of a chimpanzee in Gombe National Park, Tanzania. Folia Primatol 20:81–94. Tsukahara T 1993. Lions eat chimpanzees: the first evidence of predation by lions on wild chimpanzees. Am J Primatol 29:1–11. Uehara 1997. Predation on mammals by the chimpanzee (Pan troglodytes). Primates 38:193–214. Watts DP 2008. Scavenging by chimpanzees at Ngogo and the relevance of chimpanzee scavenging to early hominin behavioral ecology. J Hum Evol 54:125–133. Back to Contents |