|
<NOTE>
Is Chimpanzee (Pan troglodytes schweinfurthii) Low Population Density Linked with Low Levels of Aggression?
David R. Samson1 & Kevin D. Hunt2
1 Department of Evolutionary Anthropology, Duke University, USA 2 Indiana University, USA INTRODUCTION Observations have shown intraspecific aggression to be a common behavior in chimpanzee society (van Lawick-Goodall 1968; Muller 2002). Both sexes are characterized by an array of aggressive behaviors, varying in severity from non-directed displays to lethal attacks. Forces driving intragroup agonism range from male-male competition to increase status within a linear dominance hierarchy to maintaining access to estrous females (Watts 1998). Recently, Wilson et al. (2014) surveyed 18 chimpanzee intercommunity rates of lethal aggression, and showed population density to be a significant predictor. Yet, it remains unknown if this pattern can be generalized to rates of aggression within communities as well. Although intracommunity aggression is often less brutal than between community aggression, it may be driven by the same ecological forces. The goal of this study was to document the rates of aggression for the savanna-gallery forest Semliki chimpanzee (Pan troglodytes schweinfurthii) community and compare them with known rates at Kanyawara and Gombe. We test the hypothesis that population density is related to intraspecific group aggression. We predict that Semliki chimpanzees will be more peaceful than Kanyawara and Gombe chimpanzees, given Semliki has the largest of all recorded home ranges, and lowest population density among observed chimpanzee communities. METHODS Study area Chimpanzees have been studied in the Toro-Semliki Wildlife Reserve (TSWR) in western Uganda since 1996 (Samson & Hunt 2012). Their community home range is the largest known at 72.1 km2 (Samson & Hunt 2012), with the second and third largest home ranges being the dry-habitat sites of Fongoli at 64 km2 (Pruetz 2006) and Assirik at 50 km2 (Tutin et al. 1983). There are estimated to be 30 males, which suggests a community size of approximately 104 (estimated using the average sex ratio across P. t. schweinfurthii sites) (Stumpf 2007). Data collection Data were collected between the months of August 2010–January 2011. When chimpanzees were observed, we used 40-min group focal follows to generate rates of aggression for individuals (Altmann 1974). Alloccurrence sampling was possible given the conspicuous nature of chimpanzee agonism. If a party could not be observed for the full 40-min period, then the data was not used. Party composition was recorded every 10 min during focal follows. Behavioral categories followed those of Goodall (1986) and methods follow general protocols applied to characterize group and individual levels of aggression by other researchers at chimpanzee field sites (Muller 2002). Charging displays, chases and all incidents of contact aggression were considered as aggression. Data analysis All analyses were conducted in R (R Development Core Team 2014). Average rates of aggression per hour were calculated. Pearson's correlation coefficient (r) was used to assess relationships between group size, intercommunity population density (among the sites at Semliki, Gombe and Kanyawara) and agonism. A Poisson test was used for inter-site comparison. Given small sample sizes, power analysis was performed to predict ideal sample sizes required to achieve greater power properties. All statistical tests were two tailed.
RESULTS Overall, a total of 34.7 hr of observation were conducted. Semliki male chimpanzees were the only sex observed performing aggressive behaviors. Number of aggressive acts was significantly correlated with party size (r = 0.43, p = 0.001) and the average party size during an aggressive event (N = 6, mean = 6.17) was larger than the average party size when no aggressive event occurred (N = 46, mean = 3.61). Six instances of aggression were observed from the total sample (N = 52) of 40-min group focal follows. Three instances (50%) of aggression occurred under the context of reunion. Adult males at Semliki are characterized by 0.17 aggressive acts per observation hour (see Table 1). A comparison of rates by way of a Poisson test reveals that the Semliki sample was almost half as likely to show aggression than the Kanyawara sample (rate ratio = 0.56), although the result only trends towards significance. Averaged rates of aggression showed a strong, positive relationship with community population density (r2 = 0.93, p = 0.26; see Figure 1); power analysis revealed an n of 6 (sites) would achieve a power level of 0.8 and significant results at the current r2. 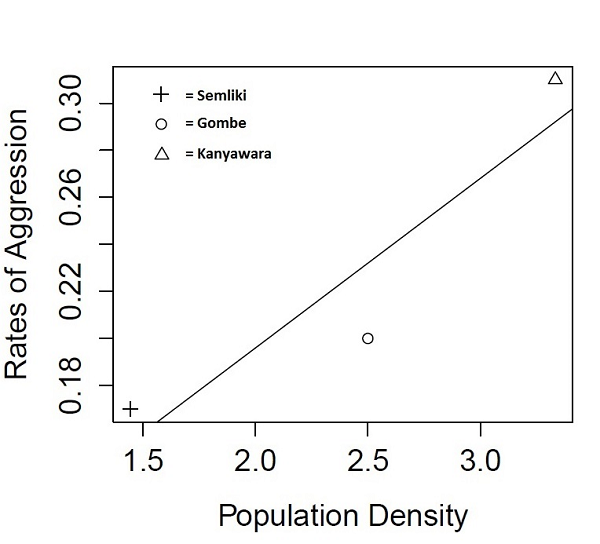
DISCUSSION To our knowledge, this study is the first to directly compare within group aggression between a large home range, low population community with that of smaller, more population dense communities. Overall, the hypothesis that population density is linked with intraspecific aggression was not rejected (due to small sample sizes further research is necessary to support the hypothesis). Male chimpanzees at Semliki seem to experience less frequent aggression (charging displays, chases and attacks) than do males at other communities (see Table 1 & Figure 1). Interesting patterns emerge from these data, which suggest that not only is intercommunity aggression a function of population density (Wilson et al. 2014), but this effect also helps explain the rates of intracommunity aggression. The adaptive benefits for intercommunity aggression has been well documented as coalitionary behavior may be an evolved tactic by which chimpanzees increase their fitness through increased access to territory, food and mates (Watts & Mitani 2001; Wilson & Wrangham 2003; Watts et al. 2006; Wrangham et al. 2006). That the pattern holds for intracommunity aggression suggests that ecology and territory size is a key predictor of violent behavior in general. Chimpanzees avoid costly encounters when possible, and a greater home-range size permits less frequent contact, and therefore less need for violent behavior. These data are preliminary, and future research should calculate rates of aggression within communities to robustly test trends suggested by this research. ACKNOWLEDGEMENTS We thank the Government of Uganda, particularly the Uganda National Council for Science and Technology and the Uganda Wildlife Authority. We thank Aggrey Rwetsiba, TSWR Warden Chris Oryema, Chief Warden Charles Tumwesigye and staff at the Semliki Wildlife Reserve for essential support. We gratefully acknowledge the National Science Foundation (SGER BNS 97-11124 and BCS 98-15991), the Semliki Chimpanzee Project, and Indiana University (Faculty Research Support Program and the College of Arts and Sciences) for financial support. We thank Holly Green for assistance in data collection. Finally, we thank Stephanie Dickinson and the Indiana Statistical Consulting Center (ISCC) provided statistical support. Finally, we thank Kazuhiko Hosaka and Michio Nakamura for thoughtful commentary on the original manuscript. REFERENCES Altmann J 1974. Observational study of behavior: Sampling methods. Behaviour 49:227–266. Chapman CA, Wrangham RW 1993. Range use of the forest chimpanzees of Kibale: Implications for the understanding of chimpanzee social organization. Am J Primatol 31:263–273. Goodall J 1986. The Chimpanzees of Gombe: Patterns of Behavior. Belknap, Cambridge, MA. Goodall JvL 1968. The behaviour of free-living chimpanzees in the Gombe Stream Reserve. Anim Behav Monogr 1:161–311. Muller MN 2002. Agonistic relations among Kanyawara chimpanzees. In: Behavioral Diversity in Chimpanzees and Bonobos. Boesch C, Hohmann G, Marchant LF (eds), Cambridge University Press, Cambridge, pp. 112–124. Pruetz JD 2006. Feeding ecology of savanna chimpanzees (Pan troglodytes verus) at Fongoli, Senegal. In: Feeding Ecology in Apes and Other Primates: Ecological, Physiological and Behavioural Aspects. Hohmann G, Robbins M, Boesch C (eds), Cambridge University Press, Cambridge, pp. 161–182. R Development Core Team 2014. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna: http:⁄⁄www.R-project.org Samson DR, Hunt KD 2012. A thermodynamic comparison of arboreal and terrestrial sleeping sites for dry-habitat chimpanzees (Pan troglodytes schweinfurthii) at the Toro-Semliki Wildlife Reserve, Uganda. Am J Primatol 74:811–818. Stumpf R 2007. Chimpanzees and bonobos: diversity within and between species. In: Primates in Perspective. Campbell CJ, Fuentes A, MacKinnon KC, Panger M, Bearder SK (eds), Oxford University Press, New York, pp. 321–344. Tutin CEG, McGrew WC, Baldwin PJ 1983. Social organization of savanna-dwelling chimpanzees, Pan troglodytes verus, at Mt. Assirik, Senegal. Primates 24:154–173. Watts DP 1998. Coalitionary mate guarding by male chimpanzees at Ngogo, Kibale National Park, Uganda. Behav Ecol Sociobiol 44:43–55. Watts DP, Mitani JC 2001. Boundary patrols and intergroup encounters in wild chimpanzees. Behaviour 138:299–327. Watts DP, Muller M, Amsler SJ, Mbabazi G, Mitani JC 2006. Lethal intergroup aggression by chimpanzees in Kibale National Park, Uganda. Am J Primatol 68:162–180. Wilson ML, Boesch C, Fruth B, et al. 2014. Lethal aggression in Pan is better explained by adaptive strategies than human impacts. Nature 513:414–418. Wilson ML, Wrangham RW 2003. Intergroup relations in chimpanzees. Ann Rev Anthropol 32:363–392. Wrangham RW, Wilson ML, Muller MN 2006. Comparative rates of violence in chimpanzees and humans. Primates 47:14–26. Back to Contents |