|
<NOTE>
Well Digging by Semliki Chimpanzees: New Data on Laterality and Possible Significance of Hydrology
William C. McGrew1,
Linda F.Marchant2,
Charlotte L.R. Payne1,
Timothy H. Webster3, & Kevin D. Hunt 4 1. Dept. of Archaeolog y & Anthropolog y, Universit y of Cambridge, UK 2. Dept. of Anthropology, Miami University, USA 3. Dept. of Anthropology, Yale University, USA 4. Dept. of Anthropology, Indiana University, USA INTRODUCTION The wild chimpanzees (Pan troglodytes schweinfurthii) of Toro–Semliki Wildlife Reserve, Uganda, are the only population of wild apes yet reported to dig wells for drinking water1. They dig holes by hand in sandy riverbeds, then drink the water collected there by mouth or with ‘sponges' made of leaves. We previously reported that the excavated tailings of this digging were often ÅebilobedÅf in distribution on the substrate, that is, clearly divided into separate piles of sand to the left and right sides of the hole (McGrew et al. 2007). Because these piles were symmetrical in volume, we inferred that the chimpanzees were manually ambilateral, that is, that they did not favour one hand or the other in digging. That is, we inferred that had their digging been right-biassed, then the rightside pile of sand would have been bigger. However, our first report was no more than a pilot study, as it was done with only 2 days (July) of intensive etho-archaeological data collection over less than 1 km of riverbed, only in the dry season. Furthermore, it was done with only a single measure, ‘volume'. Here we report a longer study done over 6 months (May–Nov.) over more than 5 km of riverbed, done in both wet and dry seasons. The aim of the follow–up study was to replicate and extend the pilot study's findings, but in the process several unexpected factors were revealed, chiefly hydrological ones. In the first study in 2006, flow of surface water in the bed of the Mugiri River had ceased, leaving only scattered, stagnant pools. In some places, the sandy substrate was still moist, but in others it was dry. In the second study in 2008, surface water flow never ceased, though it varied in depth and volume, and the sand in the riverbed was always moist. Thus, environmental affordance of drinking water for the chimpanzees showed marked interannual variation. With regard to laterality, we expected (as in the pilot study) that if the chimpanzees were manually biased at population-level to one side or another, then overall either left or right piles of tailings (lobes) would be larger. If there was no bias in the pooled data, then we expected no difference in any left– versus right–side measurements. 
Figure 1. Dry Mugiri riverbed in 2006. 
Figure 2. Mugiri riverbed in 2008. Two researchers examine well on sandbar, with flowing water behind. METHODS Full details of the study site and methods are given elsewhere (Hunt 2000; Hunt & McGrew 2002). We found 121 wells, but 67 were either too disturbed by trampling to provide data or had a single, central tailing of excavated sand, so they could not provide data on laterality. For the remaining 54 wells that were bi–lobed (that is, had distinct left and right tailings), we recorded by tape measure (cm) the following variables: well length, width and depth; length, width and depth of water in well (if present); mean distance to nearest running water; mean distance to nearest well (if present within 5 m); presence⁄absence of leaf sponges. For each lobe, we recorded: length, wide, height (which multiplied gave volume); weight (nearest 100 g, by spring balance). All variables except weight were for replication of 2006 data. Note that no data could be assigned to individuals, so data were pooled, which is usual practice for artefacts in archaeology. 
Figure 3. Measurement of water depth in well containing leaf ‘sponge'. RESULTS Despite intensive searching and monitoring over a 5–fold longer distance of riverbed and over a 6–fold longer period in 2008, we found wells at a much lower rate and had no direct observations of well digging. Thus, the study remained archaeological, that is, based on indirect evidence. Table 1 compares left vs. right lobes of bi–lobed tailings left at wells. As in 2006, there was no difference in 2008 in volume between them. Similarly, the new variable, weight, also showed no difference between left vs. right. Table 2 shows the extent of contrast between volumes of left and right lobes, that is, the ratio of R⁄L when right lobes were bigger vs. L⁄R when left lobes were bigger. In both 2006 and 2008, there was no difference between these ratios. Table 1. Left vs. right ‘lobes' of excavated sand tailings of wells compared by volume for two years; weight of lobed tailings only for 2008. 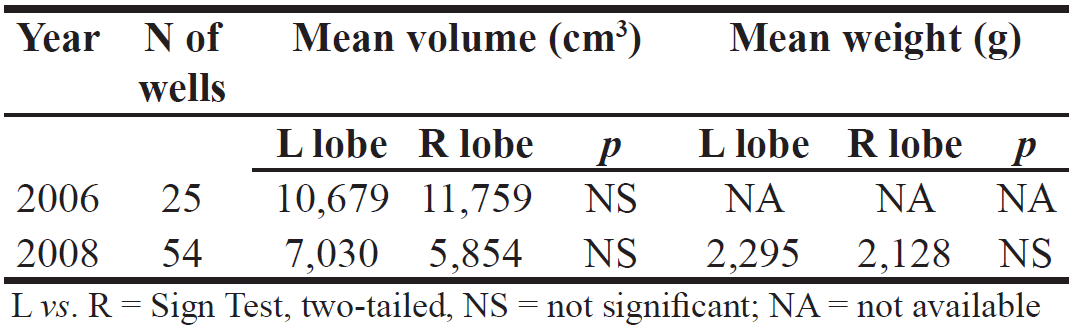
Table 2. Extent to which volumes of left vs. right tailings (lobes) (R⁄L, L⁄R) differ, for two years. 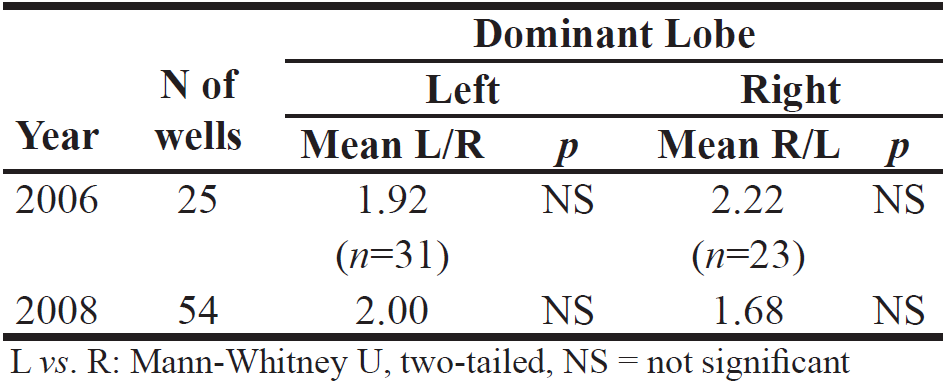
However, there were differences between years: Table 1 shows that wells were smaller in 2008: Volume of both left and right lobes was less. Table 3 shows that in 2008, only one leaf sponge tool was found at 121 wells, while in 2006, one or more sponge tools were found on average at every ninth well. Table 3. Presence or absence of leafsponge tools at well, comparing two years. 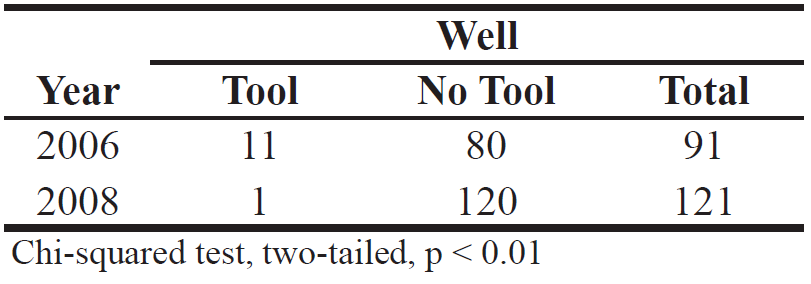
Table 4. Relationship between rainfall (mm) and abundance of wells: Month–by– month comparison of 2008 versus 1997–2006 (median). 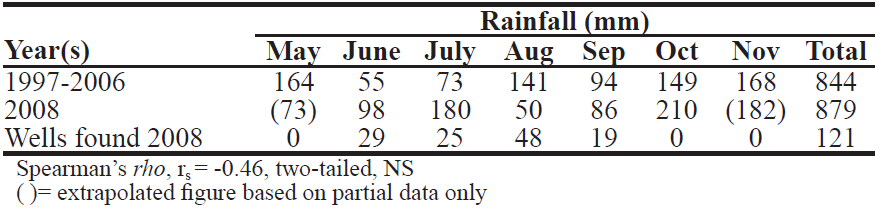
DISCUSSION The symmetry between left and right lobes was replicated across study years, adding more data to the long list of studies that have failed to find handedness in wild chimpanzees (e.g. Marchant & McGrew 2013). However, the unexpected inter–annual differences found require explanation. Overall rainfall seems not to suffice: Table 4 compares monthly rainfall totals over the study period in 2008 with average totals for a previous 10– yr period (1997–2006). Overall, 2008 was not an atypical year, although July was very wet (180 mm) and August very dry (50 mm). There was no correlation between monthly rainfall and abundance of wells. Instead, the contrast may reflect hydrological differences, that is, the level of the water table in the riverbed. A high water table allows for shallower wells and so, less sand needs to be removed, while a low water table requires deeper wells and the removal of more sand. Similarly, water can be drunk directly with the lips from a shallow well, while sponge use facilitates drinking from a deeper well. Such hydrological factors rarely, if ever, have been addressed in field studies of wild chimpanzees, but they may be crucial for explaining the presence or absence of potable drinking water in hot, dry and open habitats, especially in the dry season. Further study is needed. 
Figure 4.Leaf ‘sponges' for extracting water from wells. Top row: Intact leaves plucked from source. Middle row: Processed leaves with softer tissue removed and used but stiff veins discarded. Bottom row: Used sponges. ACKNOWLEDGEMENTS We thank: Uganda Wildlife Authority and Uganda Council for Science and Technology for permission to do research at Semliki; UWA rangers for field assistance and Semliki camp staff for domestic assistance; Phineas Hodson and Samantha Russak for research collaboration; Rebecca Jeanne Andrew Memorial Fund, Miami University (THW) for funding for field work; Leverhulme Trust (WCM) for funding for analysis and writing up results. Notes
Back to Contents |