|
<NOTE>
Chimpanzees in Bandafassi Arrondissement, Southeastern Senegal: Field Surveys as a Basis for the Sustainable Community-Based Conservation
Maja Gašperšič1 & Jill D. Pruetz2
1 Department of Biology, University of Ljubljana, Slovenia 2 Department of Anthropology, Iowa State University, USA INTRODUCTION The western chimpanzee (Pan troglodytes verus) is considered as one of the most threatened ape species, facing a dramatic decline over the last decade1,2. The latest conservation action plans classified Senegal as “an exceptionally important priority area” for chimpanzee protection, which demands immediate attention3. Chimpanzees have been expatriated from at least two African countries and IUCN estimated the Senegalese population to be almost extinct, numbering between 200 and 4004. Most apes range in small isolated communities in intense sympatry with local ethnic groups. Major threats include human encroachment, deforestation for crops, gold and iron digging, along with limited pet trade5. Additionally, this population lives at the northern edge of species’ distribution, in extremely hot, dry and open savanna landscape that characterized an important transitional period in human evolution6. Chimpanzees though have a mythical relation with Senegalese people; therefore local folklore and taboos allows them to share space. This project is part of the investigation “Conservation of chimpanzees in south-eastern Senegal: the human element” supervised by JD Pruetz. Initially, we identified ape communities in Bandafassi Arrondissement, their ranging patterns, key water and food sources, and particularly chimpanzees’ relation to humans via an ethnoprimatological approach7. Due to the importance of water sources with gallery forests, specific food-rich areas, and conflicts with humans encountered we focused on three priority field-sites: mountain slopes above Bandafassi village, where chimpanzees were known to attack goats; the stream of Angafou (12°34′N, 12°24′W), its rich gallery forest and mango orchards, being crop-raided by apes, supposedly after commercial harvesting of baobab and Saba fruits increased; and the surroundings of Nathia (12°29′N, 12°22′W), which was not identified in previous surveys2 and shows no conflict between species. Two buffer-zones are surveyed sporadically to provide basic information about the presence of chimpanzees in the remaining forest patches between the Niokolo Koba national park and neighboring Guinea-Conakry. Several sites along Gambia River indicate various conflicts between chimpanzees and humans (e.g. palm-wine harvesting, raphia or bamboo-cutting, artisanal gold-digging sites). Chimpanzees are seen there only when people are not common, and it appears that their seasonal activity affects the ranging patterns of apes. METHODS Habitat composition at study sites was sampled by walking several km in random directions and assessing the general habitat within a 50 m radius each 30 m. The classification of vegetation types was adapted after Bogart & Pruetz8. In March 2011 a phenology transect (1 km long, 10 m wide) was established at each study site to monitor tree production monthly throughout the year. Chimpanzee and human usage is assessed here on a total of 507 woody plants. Key food sources (e.g. Adansonia digitata, Cola cordifolia, Parkia biglobosa, etc.), and termite mounds and beehives were marked as well. We also noted encounters with large mammals and humans to estimate the presence of predators, competitors and potential prey-species for chimpanzees. Data were systematically collected upon sightings, nesting9 and feeding sites, while signs of material culture10 were recorded opportunistically. Diet composition was assessed through observations, feeding traces, and fecal-sample analyses11. Identifying ranging behavior included recording signs of chimpanzee activity and, if indices were positive (fresh debris or vocalizations), we tried to approach them. RESULTS From April 2010 through June 2011 we conducted 292 field surveys and identified 13 villages along several dry season water sources associated with chimpanzees. In total we recorded 157 nesting sites, 85 feeding sites, several cases of tool-use (e.g. baobab cracking, termite-fishing, ant-dipping, tuber-digging), hand-dug wells and caves/shelters. Chimpanzees of Bandafassi were encountered more than 160 times, while data on demography, activity budget and reactions to observers stems from 146 observations at three study sites. Encounters with chimpanzees Despite the rarity of closed-canopy habitat12, chimpanzees were most often observed in gallery and ecotone forest (38.3% and 0.8% respectively; see Figure 1), followed by different woodland types (closed 26.3%, open 19.1% and bamboo 9.2%), on plateaus (3.5%), in grassland and along fields (1.4% both types). These results indicate chimpanzees’ preference for forested habitats, especially for nesting. This habitat type is also most heavily exploited by humans, particularly nomadic pastoralists13. 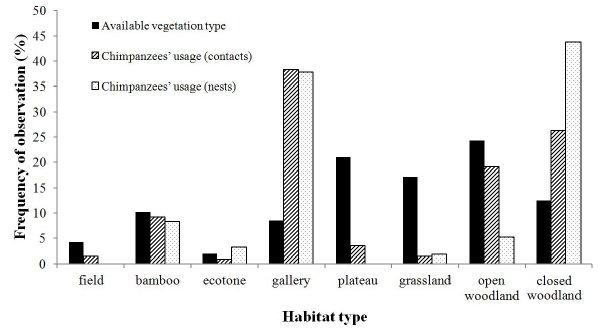
Figure 1. Habitat composition/available vegetation types compared with chimpanzees’ usage (nests, contacts) The average party size for various chimpanzee groups sighted was 5.8 (range 1–30). The largest daily parties were encountered at Nathia (mean 7.8), followed by Angafou (5.6), while the smallest groups ranged at Bandafassi (4.5). Foraging parties were on average composed of 2.4 males, 2.1 females, 1.6 juveniles, and 1.3 infants. The community at Angafou regularly nests along the stream and has indicated some acceptance of observers through time, while Nathia chimpanzees seem to have a vast home-range (> 70 km2) and use it seasonally. As expected, apes were most often feeding (63.4% overall) upon sighting, foraging and traveling (17.9%), drinking (8.3%) resting (4.8%), nesting (2.1%) and other (in 3.4% mostly social behaviors). The most common reaction of chimpanzees in general was to leave the location (38% overall; see Figure 2), but positively they ignored us and indicated curiosity more often over time (24% and 20% respectively). However, important differences in reactions occurred among three sites with different human-chimpanzee relations. The apes indicated fear in only 14.3% of all episodes, but more significantly at Angafou. Nathia chimpanzees exhibited less fear than other communities and often ignored observers during prolonged contacts, while only apes at Bandafassi also displayed at human observers (4.3%). 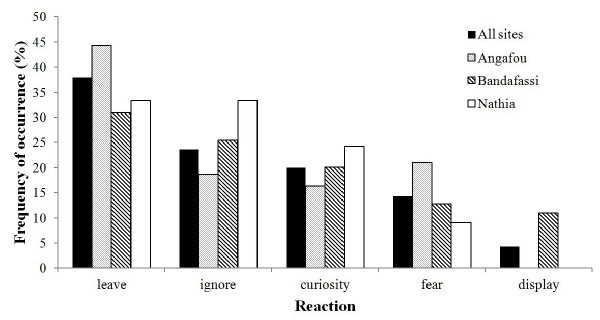
Figure 2. Comparison of chimpanzee reactions to humans among three main sites Competition with humans So far, chimpanzees consumed fruits (73.2%), pods (7.6%), nuts (4.9%), pith (4.9%), bark (2.9%), leaves (1.5%; partly used for medicinal purposes) and flowers (0.4%) from 39 plant species of at least 17 families. Social insects (termites and weaver ants together in 2.6%), tubers (wild yam and unidentified underground storage organs), honey (each in 0.4% of cases) and traces of geophagy were detected in their food repertoire. Many ape food sources are also used by local humans for various purposes (Pruetz listed 17 species14), and at least five of their key feeding items are exploited commercially (e.g. Adansonia digitata, Saba senegalensis, Tamarindus indica, Parkia biglobosa, Cola cordifolia). Among tree species used for food or nest construction several were also cut by shepherds; e.g. Bombax costatum, Khaya senegalensis, Acacia spp., Ficus spp., and Zizyphus spp.13. Fulbe herders from North of the country were in fact most often encountered in the field, pruning trees for their large flocks of sheep (100–300), while harvesting of Saba fruits occurred as a key activity by local people in June. DISCUSSION: A Vision For The Sustainable Community-Based Conservation Chimpanzees’ responses to observers appear to be influenced by their experience with local people, who are the crucial element for their conservation15. Nature-culture tourism programs, including forest guards, could be an alternative solution to the complex relationship between the species16, while habituating the apes is not recommended17. The incentive could include local traditional customs related to primates (Bedik and Bassari initiation rites) and nature hiking with the opportunistic chance of chimpanzee viewing. This particular research project covers the widest geographical area in Senegal (> 500 km2), including several chimpanzee communities in rare forest patches. As of December 2011, five field assistants, serving as eco-rangers, monitor the most important chimpanzee sites. We established a limited “surveillance system” similar to J. Carter’s in Guinea or further east in Senegal. Identified eco-guards monitor the ranging behavior of chimpanzees and at the same time limit crop-raiding and attacks on domestic animals. The two bordering-zones are important in terms of recognizing the existing forest-corridors available for possible gene-transfer within the population. It appears that with the continuous presence of researchers the apes ceased to capture goats, similarly as mango-raiding reportedly decreased. Encounters with shepherds, wine-collectors or hunters in the field are reported to local Eaux et forets authorities. Preliminary estimates from surveys in Senegal by S Ndiaye in May 2011 are encouraging and indicate the Senegalese population is stronger than thought and numbers up to 500 chimpanzees18. In fact, two of our main study areas (Angafou and Nathia) were recognized as priority sites for chimpanzee conservation18. To conclude, this project has laid solid foundations for the sustainable community-based conservation in Bandafassi arrondissement. However, the prospects for the future co-existence of savanna chimpanzees and humans depend on needed support from governmental agencies as well as investment in terms of research and funding. Acknowledgements First we want to thank Senegalese department of Eaux et forêts at the Ministry of environment and the Arrondissement du Bandafassi to give us the permission for research. The project was generously supported by the Slovenian Research Agency ARRS (Z6-3676). We are sincerely grateful to local authorities and villagers for accepting our presence in their forests, USAID organization Wula Nafaa for initial funding of the project, Fongoli savanna chimpanzee project for logistics and Dondo Kanté for partly managing the project. Collaboration with Janis Carter and Soulèye Ndiaye was invaluable. The research though would not be possible without dedicated field assistants / forest guardians: Gilbert Camara at Bandafassi mountains, Nambura Diallo at Nathia, Khadim Diallo at park borders, Samba Faye Kanté at Angafou, and Omar Diallo at Marewa (along with various tasks). Finally, special thanks goes to Seth Cissé for his overall help, data-collection and supervising the project in my absence. REFERENCES
Back to Contents |